
A Novel Strain of Tomato Leaf Curl New Delhi Virus Has Spread to the Mediterranean Basin

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Sampling
2.2. Sample Analysis and Genetic Diversity Study
2.3. Phylogenetic Analysis
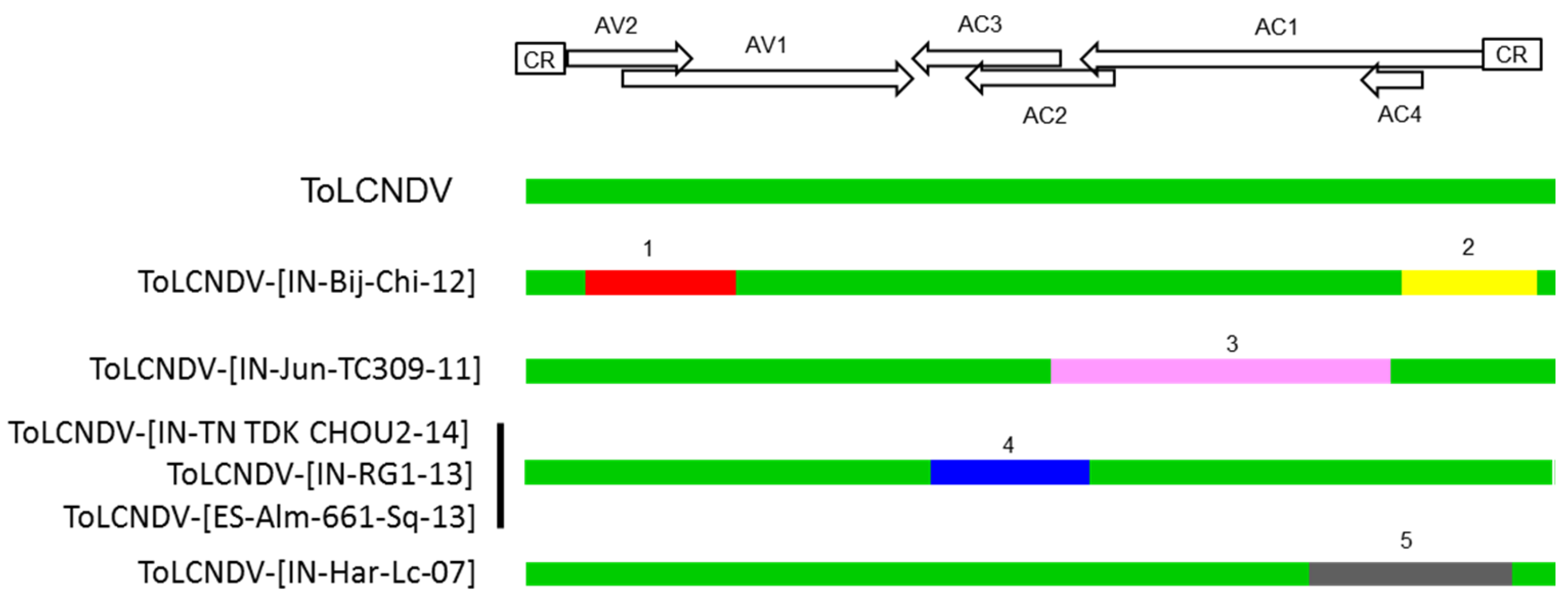
2.4. Recombination Analysis
2.5. Construction of Infectious Clones and Agroinoculation
2.6. Host Range Study
2.7. Whitefly Transmission
3. Results
3.1. Widespread Occurrence of ToLCNDV in Spanish Vegetable Growing Regions: Evidence for a Genetically-Uniform Population
3.2. Genetic Relationships of ToLCNDV Isolates: A Complex Group with a New Strain of Recombinant Nature Emerging in Spain
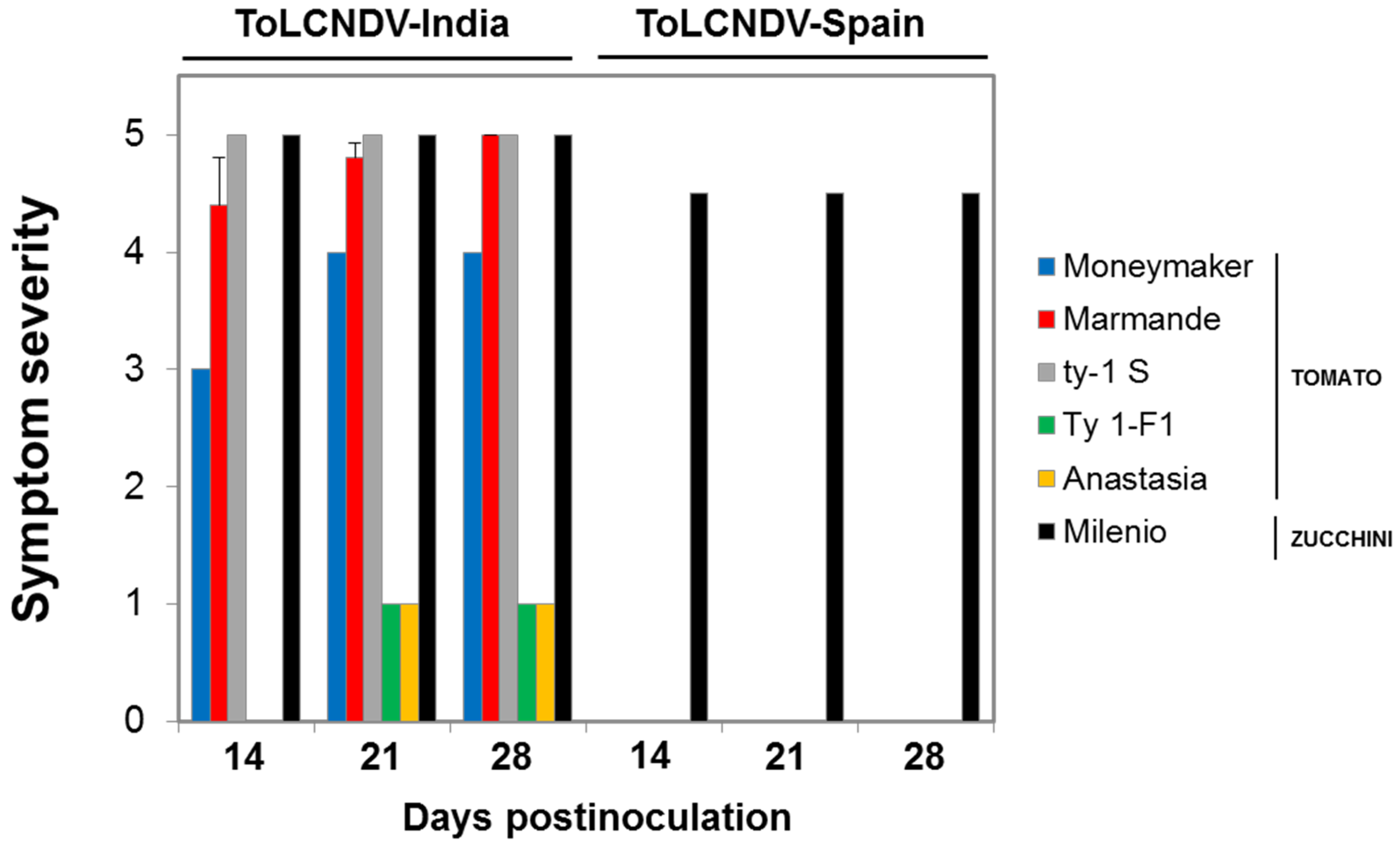
3.3. The ToLCNDV Isolates of the Novel Strain Emerging in Spain Are Infectious in Cucurbits, but Poorly Adapted to Tomato
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Navas-Castillo, J.; Fiallo-Olivé, E.; Sánchez-Campos, S. Emerging virus diseases transmitted by whiteflies. Annu. Rev. Phytopathol. 2011, 49, 219–248. [Google Scholar] [CrossRef] [PubMed]
- Varma, A.; Mandal, B.; Singh, M. Global emergence and spread of whitefly (Bemisia tabaci) transmitted geminiviruses. In The Whitefly, Bemisia tabaci (Homoptera: Aleyrodidae) Interaction with Geminivirus-Infected Host Plants; Thompson, W.M.O., Ed.; Springer: Dordrecht, The Netherlands, 2011; pp. 205–292. [Google Scholar]
- Brown, J.; Zerbini, F.M.; Navas-Castillo, J.; Moriones, E.; Ramos-Sobrinho, R.; Silva, J.C.; Fiallo-Olivé, E.; Briddon, R.; Hernández-Zepeda, C.; Idris, A.; et al. Revision of Begomovirus taxonomy based on pairwise sequence comparisons. Arch. Virol. 2015, 160, 1593–1619. [Google Scholar] [CrossRef] [PubMed]
- Varsani, A.; Navas-Castillo, J.; Moriones, E.; Hernández-Zepeda, C.; Idris, A.; Brown, J.; Murilo Zerbini, F.; Martin, D. Establishment of three new genera in the family Geminiviridae: Becurtovirus, Eragrovirus and Turncurtovirus. Arch. Virol. 2014, 159, 2193–2203. [Google Scholar] [CrossRef] [PubMed]
- Padidam, M.; Beachy, R.N.; Fauquet, C.M. Tomato leaf curl geminivirus from India has a bipartite genome and coat protein is not essential for infectivity. J. Gen. Virol. 1995, 76, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Mansoor, S.; Khan, S.H.; Saeed, M.; Bashir, A.; Zafar, Y.; Malik, K.A. Evidence for the association of a bipartite geminivirus with tomato leaf curl disease in Pakistan. Plant Dis. 1997, 81. [Google Scholar] [CrossRef]
- Usharani, K.S.; Surendranath, B.; Paul-Khurana, S.M.; Garg, I.D.; Malathi, V.G. Potato leaf curl—A new disease of potato in northern India caused by a strain of Tomato leaf curl New Delhi virus. Plant Pathol. 2004. [Google Scholar] [CrossRef]
- Hussain, M.; Mansoor, S.; Iram, S.; Zafar, Y.; Briddon, R.W. First report of Tomato leaf curl New Delhi virus affecting chilli pepper in Pakistan. Plant Pathol. 2004. [Google Scholar] [CrossRef]
- Pratap, D.; Kashikar, A.; Mukherjee, S. Molecular characterization and infectivity of a Tomato leaf curl New Delhi virus variant associated with newly emerging yellow mosaic disease of eggplant in India. Virol. J. 2011, 8. [Google Scholar] [CrossRef] [PubMed]
- Sohrab, S.S.; Mandal, B.; Pant, R.P.; Varma, A. First report of association of Tomato leaf curl virus-New Delhi with yellow mosaic disease of Luffa cylindrica in India. Plant Dis. 2003, 87. [Google Scholar] [CrossRef]
- Ito, T.; Sharma, P.; Kittipakorn, K.; Ikegami, M. Complete nucleotide sequence of a new isolate of Tomato leaf curl New Delhi virus infecting cucumber, bottle gourd and muskmelon in Thailand. Arch. Virol. 2008, 153, 611–613. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Mishra, K.K.; Chattopadhyay, B.; Chakraborty, S. Biological and molecular characterization of a begomovirus associated with yellow mosaic vein mosaic disease of pumpkin from Northern India. Virus Genes 2009, 39, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.H.; Ku, H.M.; Tsai, W.S.; Chien, R.C.; Jan, F.J. Identification and characterization of a mechanical transmissible begomovirus causing leaf curl on oriental melon. Eur. J. Plant Pathol. 2010, 127, 219–228. [Google Scholar] [CrossRef]
- Mizutani, T.; Daryono, B.S.; Ikegami, M.; Natsuaki, K.T. First report of Tomato leaf curl New Delhi virus infecting cucumber in central Java, Indonesia. Plant Dis. 2011, 95. [Google Scholar] [CrossRef]
- Chakraborty, S. Tomato leaf curl viruses from India (Geminiviridae). In Enciclopedy of Virology; Mahy, B.W.J., van Regenmortel, M.H.V., Eds.; Elsevier: Amsterdam, The Netherlands, 2008; pp. 124–133. [Google Scholar]
- Zaidi, S.S.-A.; Martin, D.P.; Amin, I.; Farooq, M.; Mansoor, S. Tomato leaf curl New Delhi virus; a widespread bipartite begomovirus in the territory of monopartite begomoviruses. Mol. Plant Pathol. 2016, in press. [Google Scholar] [CrossRef] [PubMed]
- Juárez, M.; Tovar, R.; Fiallo-Olivé, E.; Aranda, M.A.; Gosálvez, B.; Castillo, P.; Moriones, E.; Navas-Castillo, J. First detection of Tomato leaf curl New Delhi virus infecting zucchini squash in Spain. Plant Dis. 2014, 98. [Google Scholar] [CrossRef]
- López, C.; Ferriol, M.; Picó, M. Mechanical transmission of Tomato leaf curl New Delhi virus to cucurbit germplasm: Selection of tolerance sources in Cucumis melo. Euphytica 2015, 204, 679–691. [Google Scholar] [CrossRef]
- Ruiz, M.L.; Simón, A.; Velasco, L.; García, M.C.; Janssen, D. First report of Tomato leaf curl New Delhi virus infecting tomato in Spain. Plant Dis. 2015, 99. [Google Scholar] [CrossRef]
- Mnari-Hattab, M.; Zammouri, S.; Belkadhi, M.S.; Bellon Doña, D.; ben Nahia, E.; Hajlaoui, M.R. First report of Tomato leaf curl New Delhi virus infecting cucurbits in Tunisia. New Dis. Rep. 2015, 31. [Google Scholar] [CrossRef]
- Panno, S.; Lacono, G.; Davino, M.; Marchione, S.; Zappardo, V.; Bella, P.; Tomassoli, L.; Accotto, G.P.; Davino, S. First report of Tomato leaf curl New Delhi virus affecting zucchini squash in an important horticultural area of southern Italy. New Dis. Rep. 2016, 33. [Google Scholar] [CrossRef]
- Sáez, C.; Martínez, C.; Ferriol, M.; Manzano, S.; Velasco, L.; Jamilena, M.; Lopez, C.; Picó, B. Resistance to Tomato leaf curl New Delhi virus in Cucurbita spp. Ann. Appl. Biol. 2016, 169, 91–105. [Google Scholar] [CrossRef]
- Ji, Y.; Scott, J.; Hanson, P.; Graham, E.; Maxwell, D. Sources of resistance, inheritance, and location of genetic loci conferring resistance to members of the tomato-infecting begomoviruses. In Tomato Yellow Leaf Curl Virus Disease; Czosnek, H., Ed.; Springer: Dordrecht, The Netherlands, 2007; pp. 343–362. [Google Scholar]
- Navas-Castillo, J.; Sánchez-Campos, S.; Díaz, J.A.; Sáez-Alonso, E.; Moriones, E. Tomato yellow leaf curl virus-Is causes a novel disease of common bean and severe epidemics in tomato in Spain. Plant Dis. 1999, 83, 29–32. [Google Scholar] [CrossRef]
- García-Andrés, S.; Monci, F.; Navas-Castillo, J.; Moriones, E. Begomovirus genetic diversity in the native plant reservoir Solanum nigrum: Evidence for the presence of a new virus species of recombinant nature. Virology 2006, 350, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Haible, D.; Kober, S.; Jeske, H. Rolling circle amplification revolutionizes diagnosis and genomics of geminiviruses. J. Virol. Methods 2006, 135, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Inoue-Nagata, A.K.; Albuquerque, L.C.; Rocha, W.B.; Nagata, T. A simple method for cloning the complete begomovirus genome using the bacteriophage φ29 DNA polymerase. J. Virol. Methods 2004, 116, 209–211. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucl. Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Muhire, B.M.; Varsani, A.; Martin, D.P. SDT: A virus classification tool based on pairwise sequence alignment and identity calculation. PLoS ONE 2014, 9, e108277. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kumar, R.V.; Sharma, V.K.; Chattopadhyay, B.; Chakraborty, S. An improved plant regeneration and Agrobacterium—Mediated transformation of red pepper (Capsicum annuum L.). Physiol. Mol. Biol. Plants 2012, 18, 357–364. [Google Scholar] [CrossRef] [PubMed]
- De Castro, A.P.; Blanca, J.M.; Díaz, M.J.; Nuez, F. Identification of a CAPS marker tightly linked to the Tomato yellow leaf curl disease resistance gene Ty-1 in tomato. Eur. J. Plant Pathol. 2007, 117, 347–356. [Google Scholar] [CrossRef]
- De Barro, P.J.; Liu, S.S.; Boykin, L.M.; Dinsdale, A.B. Bemisia tabaci: A statement of species status. Annu. Rev. Entomol. 2011, 56, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Gilbertson, R.L.; Batuman, O.; Webster, C.G.; Adkins, S. Role of the insect supervectors Bemisia tabaci and Frankliniella occidentalis in the emergence and global spread of plant viruses. Annu. Rev. Virol. 2015, 2, 67–93. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.A.C.; Kehoe, M.A. A proposal to rationalize within-species plant virus nomenclature: Benefits and implications of inaction. Arch. Virol. 2016, 161, 2051–2057. [Google Scholar] [CrossRef] [PubMed]
- Jyothsna, P.; Haq, Q.M.I.; Singh, P.; Sumiya, K.V.; Praveen, S.; Rawat, R.; Briddon, R.W.; Malathi, V.G. Infection of Tomato leaf curl New Delhi virus (ToLCNDV), a bipartite begomovirus with betasatellites, results in enhanced level of helper virus components and antagonistic interaction between DNA B and betasatellites. Appl. Microbiol. Biotechnol. 2013, 97, 5457–5471. [Google Scholar] [CrossRef] [PubMed]
- Lefeuvre, P.; Moriones, E. Recombination as a motor of host switches and virus emergence: Geminiviruses as case studies. Curr. Opin. Virol. 2015, 10, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Seal, S.E.; van den Bosch, F.; Jeger, M.J. Factors influencing begomovirus evolution and their increasing global significance: Implications for sustainable control. Crit. Rev. Plant Sci. 2006, 25, 23–46. [Google Scholar] [CrossRef]
- Monci, F.; Sánchez-Campos, S.; Navas-Castillo, J.; Moriones, E. A natural recombinant between the geminiviruses Tomato yellow leaf curl Sardinia virus and Tomato yellow leaf curl virus exhibits a novel pathogenic phenotype and is becoming prevalent in Spanish populations. Virology 2002, 303, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.P.; Liu, Y.L.; Calvert, L.; Munoz, C.; Otim-Nape, G.W.; Robinson, D.J.; Harrison, B.D. Evidence that DNA-A of a geminivirus associated with severe cassava mosaic disease in Uganda has arisen by interspecific recombination. J. Gen. Virol. 1997, 78, 2101–2111. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, L.; Simon, A.; Velasco, L.; Janssen, D. Biological characterization of Tomato leaf curl New Delhi virus from Spain. Plant Pathol. 2016, in press. [Google Scholar] [CrossRef]
- Fraile, A.; Malpica, J.M.; Aranda, M.A.; Rodríguez-Cerezo, E.; García-Arenal, F. Genetic diversity in tobacco mild green mosaic tobamovirus infecting Nicotiana glauca. Virology 1996, 223, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Novella, I.S.; Elena, S.F.; Moya, A.; Domingo, E.; Holland, J.J. Size of genetic bottlenecks leading to virus fitness loss is determined by mean initial population fitness. J. Virol. 1995, 69, 2869–2872. [Google Scholar] [PubMed]
- Sánchez-Campos, S.; Díaz, J.A.; Monci, F.; Bejarano, E.R.; Reina, J.; Navas-Castillo, J.; Aranda, M.A.; Moriones, E. High genetic stability of the begomovirus Tomato yellow leaf curl Sardinia virus in southern Spain over an 8-year period. Phytopathology 2002, 92, 842–849. [Google Scholar] [CrossRef] [PubMed]
- Duff-Farrier, C.R.A.; Bailey, A.M.; Boonham, N.; Foster, G.D. A pathogenicity determinant maps to the N-terminal coat protein region of the Pepino mosaic virus genome. Mol. Plant. Pathol. 2015, 16, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Salvador, B.; Sáenz, P.; Yanguez, E.; Quiot, J.B.; Quiot, L.; Delgadillo, M.O.; García, J.A.; Simón-Mateo, C. Host-specific effect of P1 exchange between two potyviruses. Mol. Plant. Pathol. 2008, 9, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Michelson, I.; Zamir, D.; Czosnek, H. Accumulation and translocation of tomato yellow leaf curl virus (TYLCV) in a Lycopersicon esculentum breeding line containing the L. chilense TYLCV tolerance gene Ty-1. Phytopathology 1994, 84, 928–933. [Google Scholar] [CrossRef]
- Kumar, A.; Tiwari, K.L.; Datta, D.; Singh, M. Marker assisted gene pyramiding for enhanced Tomato leaf curl virus disease resistance in tomato cultivars. Biol. Plant 2014, 58, 792–797. [Google Scholar] [CrossRef]
- Prasanna, H.C.; Sinha, D.P.; Rai, G.K.; Krishna, R.; Kashyap, S.P.; Singh, N.K.; Singh, M.; Malathi, V.G. Pyramiding Ty-2 and Ty-3 genes for resistance to monopartite and bipartite tomato leaf curl viruses of India. Plant Pathol. 2015, 64, 256–264. [Google Scholar] [CrossRef]
- Moriones, E.; Navas-Castillo, J. Tomato yellow leaf curl disease epidemics. In Bemisia: Bionomics and Management of a Global Pest; Stansly, P.A., Naranjo, S.E., Eds.; Springer: Dordrecht, The Netherlands; Heidelberg, Germany; London, UK; New York, NY, USA, 2010; pp. 259–282. [Google Scholar]
- Anfoka, G.; Haj Ahmad, F.; Abhary, M.; Hussein, A. Detection and molecular characterization of viruses associated with tomato yellow leaf curl disease in cucurbit crops in Jordan. Plant Pathol. 2009, 58, 754–762. [Google Scholar] [CrossRef]
- Paz-Carrasco, L.; Castillo-Urquiza, G.; Lima, A.; Xavier, C.; Vivas-Vivas, L.; Mizubuti, E.; Zerbini, F.M. Begomovirus diversity in tomato crops and weeds in Ecuador and the detection of a recombinant isolate of rhynchosia golden mosaic Yucatan virus infecting tomato. Arch. Virol. 2014, 159, 2127–2132. [Google Scholar] [CrossRef] [PubMed]
- Pita, J.S.; Fondong, V.N.; Sangare, A.; Otim-Nape, G.W.; Ogwal, S.; Fauquet, C.M. Recombination, pseudorecombination and synergism of geminiviruses are determinant keys to the epidemic of severe cassava mosaic disease in Uganda. J. Gen. Virol. 2001, 82, 655–665. [Google Scholar] [CrossRef] [PubMed]
- Tiendrébéogo, F.; Lefeuvre, P.; Hoareau, M.; Harimalala, M.A.; de Bruyn, A.; Villemot, J.; Traoré, V.S.E.; Konaté, G.; Traoré, A.S.; Barro, N.; et al. Evolution of African cassava mosaic virus by recombination between bipartite and monopartite begomoviruses. Virol. J. 2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.K.; Kushwaha, N.; Chakraborty, S. Synergistic interaction among begomoviruses leads to the suppression of host defense-related gene expression and breakdown of resistance in chilli. Appl. Microbiol. Biotechnol. 2016, 100, 4035–4049. [Google Scholar] [CrossRef] [PubMed]
- Saunders, K.; Salim, N.; Mali, V.R.; Malathi, V.G.; Briddon, R.; Markham, P.G.; Stanley, J. Characterisation of Sri Lankan cassava mosaic virus and Indian cassava mosaic virus: Evidence for acquisition of a DNA B component by a monopartite Begomovirus. Virology 2002, 293, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Sanderfoot, A.A.; Lazarowitz, S.G. Getting it together in plant virus movement: Cooperative interactions between bipartite geminivirus movement proteins. Trends Cell Biol. 1996, 6, 353–358. [Google Scholar] [CrossRef]
- Alves-Júnior, M.; Alfenas-Zerbini, P.; Andrade, E.C.; Esposito, D.A.; Silva, F.N.; da Cruz, A.C.F.; Ventrella, M.C.; Otoni, W.C.; Zerbini, F.M. Synergism and negative interference during co-infection of tomato and Nicotiana benthamiana with two bipartite begomoviruses. Virology 2009, 387, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Fondong, V.N.; Pita, J.S.; Rey, M.E.C.; de Kochko, A.; Beachy, R.N.; Fauquet, C.M. Evidence of synergism between African cassava mosaic virus and a new double-recombinant geminivirus infecting cassava in Cameroon. J. Gen. Virol. 2000, 81, 287–297. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Province | No. of ToLCNDV Positive Samples/No. of Total Analyzed Samples/No. of Fields Surveyed a | ||
|---|---|---|---|---|
| Zucchini Squash | Melon | Tomato | ||
| 2013 | Málaga | 6/6/2 | - | - |
| Almería | 20/20/3 | - | - | |
| Murcia | 4/4/2 | - | - | |
| 2014 | Málaga | 8/8/1 | 1/1/1 | - |
| Almería | 24/26/3 | 2/2/1 | 0/3/2 | |
| Murcia | - | - | 0/10/1 | |
| 2015 | Málaga | 10/10/1 | 6/6/1 | 0/81/3 |
| Almería | - | - | 0/17/2 | |
| Murcia | - | - | 0/1/1 | |
| Total | 72/74/12 | 9/9/3 | 0/112/9 | |
| Event Number | Recombinant Sequence | Putative Non-ToLCNDV Parent | Putative ToLCNDV Parent | Breakpoints a | Methods b | p-Value c | |
|---|---|---|---|---|---|---|---|
| Begin | End | ||||||
| 1 | ToLCNDV-[IN-Bij-Chi-12] | ChiLCPKV-PK[PK-Kha-04] | ToLCNDV-[BG-Jes-Svr-05] | 146 | 547 | RGBMCST | 8642 × 10−16 |
| 2 | ToLCNDV-[IN-Bij-Chi-12] | Unknown d | ToLCNDV-[IN-ND-Pap-05] | 2328 | 2743 | RGBMCST | 4332 × 10−15 |
| 3 | ToLCNDV-[IN-Jun-TC309-11] | ToLCPalV-[IN-Rau-TC238-09] | ToLCNDV-[IN-ND-Pap-05] | 1397 | 2301 | RGBMCST | 4722 × 10−7 |
| 4 | ToLCNDV-[IN-TN TDK CHOU2-14] | ToLCPalV-[IR-Jir-T69P-Cuc-08] | ToLCNDV-[IN-Har-Lc-07] | 1055 | 1532 | RMC | 8702 × 10−5 |
| 4 | ToLCNDV-[IN-RG1-13] | ToLCPalV-[IR-Jir-T69P-Cuc-08] | ToLCNDV-[IN-Har-Lc-07] | 1054 | 1524 | RMC | 8702 × 10−5 |
| 5 | ToLCNDV-[IN-Har-Lc-07] | Unknown | ToLCNDV-[IN-ND-Pap-05] | 2063 | 2661 | RGBMCST | 6679 × 10−11 |
| 4 | ToLCNDV-[ES-Alm-661-Sq-13] | ToLCPalV-[IR-Jir-T69P-Cuc-08] | ToLCNDV-[IN-Har-Lc-07] | 1054 | 1531 | RMC | 8702 × 10−5 |
| Host Type | Family | Species | Cultivar/Genotype | Infected Plants/Total Inoculated Plants a | ||
|---|---|---|---|---|---|---|
| Spanish Isolate b | Indian Isolate c | |||||
| Assay 1 | Assay 2 | Assay 1 | ||||
| Cultivated host | Cucurbitaceae | Zucchini (Cucurbita pepo) | Milenio | 5/5 | 5/5 | 5/5 |
| Afrodita | 5/5 | 5/5 | - | |||
| Melon (Cucumis melo) | C-278 | 3/5 | 0/5 | - | ||
| Watermelon (Citrullus lanatus) | SV80287 | 4/5 | 0/5 | - | ||
| Cucumber (Cucumis sativus) | Pacer | 5/5 | 0/5 | - | ||
| Bell Puig | 4/5 | 2/5 | - | |||
| Fabaceae | Bean (Phaseolus vulgaris) | Donna | 0/5 | 0/5 | - | |
| Solanaceae | Tomato (Solanum lycopersicum) | Moneymaker | 0/10 | 1/10 | 9/10 | |
| Pepper (Capsicum annuum) | Jalapeño | 0/5 | 0/5 | - | ||
| C. Wonder | 0/5 | 0/5 | - | |||
| Experimental host | Solanaceae | Nicotiana benthamiana | 5/5 | 5/5 | 5/5 | |
| Nicotiana tabacum | Xanthi | 3/5 | 1/5 | - | ||
| Nicotiana glutinosa | 5/5 | 5/5 | - | |||
| Wild host | Solanaceae | Solanum nigrum | 0/5 | 0/5 | - | |
| Crop/Cultivar or Line | Agroinoculation | Whitefly Inoculation | ||||||
|---|---|---|---|---|---|---|---|---|
| Zucchini Source a | Tomato Source a | |||||||
| Spanish Isolate b | Indian Isolate d | Spanish Isolate b | Spanish Field Inoculum c | Spanish Isolate b | ||||
| MED Species | MED Species | MEAM1 Species | MED Species | |||||
| Infected Plants/Total Plants Inoculated | Infected Plants/Total Plants Inoculated | Infected Plants/Total Plants Inoculated | Infected Plants/Total Plants Inoculated | Infected Plants/Total Plants Inoculated | Infected Plants/Total Plants Inoculated | |||
| Assay 1 | Assay 2 | Assay 3 | Assay 1 | |||||
| TOMATO | ||||||||
| Moneymaker | 1/10 | - | 0/10 | 10/10 | 1/10 | 0/10 | 0/10 | 0/10 |
| Fortuna C | 2/10 | 1/10 | - | 7/10 | - | - | - | - |
| Gardener’s Delight | 0/10 | 0/10 | - | 10/10 | - | - | - | - |
| Rondeño | 2/10 | 2/10 | 1/10 | 10/10 | 0/10 | 0/10 | 0/10 | 0/10 |
| Marmande | 4/10 | 1/10 | - | 10/10 | 0/10 | 0/10 | 0/10 | 0/10 |
| Ty-1 S | 1/10 | 0/10 | - | 10/10 | 0/10 | 1/10 | 0/10 | - |
| Ty-1 F1 | 0/10 | 0/10 | - | 10/10 e | 0/10 | 0/10 | 0/10 | - |
| Anastasia | 0/10 | 0/10 | - | 2/10 e,f | - | - | - | - |
| ZUCCHINI | - | |||||||
| Milenio | 5/5 | 5/5 | 5/5 | 5/5 | 5/5 | 5/5 | 5/5 | 5/5 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fortes, I.M.; Sánchez-Campos, S.; Fiallo-Olivé, E.; Díaz-Pendón, J.A.; Navas-Castillo, J.; Moriones, E. A Novel Strain of Tomato Leaf Curl New Delhi Virus Has Spread to the Mediterranean Basin. Viruses 2016, 8, 307. https://doi.org/10.3390/v8110307
Fortes IM, Sánchez-Campos S, Fiallo-Olivé E, Díaz-Pendón JA, Navas-Castillo J, Moriones E. A Novel Strain of Tomato Leaf Curl New Delhi Virus Has Spread to the Mediterranean Basin. Viruses. 2016; 8(11):307. https://doi.org/10.3390/v8110307
Chicago/Turabian StyleFortes, Isabel M., Sonia Sánchez-Campos, Elvira Fiallo-Olivé, Juan A. Díaz-Pendón, Jesús Navas-Castillo, and Enrique Moriones. 2016. "A Novel Strain of Tomato Leaf Curl New Delhi Virus Has Spread to the Mediterranean Basin" Viruses 8, no. 11: 307. https://doi.org/10.3390/v8110307