


Abstract
Background: Bortezomib is an anti-neoplastic drug acting against multiple myeloma but its use is associated with the onset of painful peripheral neuropathy. Tumor necrosis factor-α (TNFα) is associated with the development of neuropathic pain; several models have shown that the inactivation of TNFα leads to a reduction in pain stimuli perception. The aim of the present study was to analyze if the administration of an antibody against TNFα is able to prevent the development of bortezomib-induced neuropathic pain. Materials and Methods: Nerve conduction velocity was measured and a histopathological examination was performed to assess the extent of peripheral neuropathy. To study the onset of painful neuropathy, the response to mechanical or thermal stimuli was evaluated. Results: This study demonstrated that co-administration of an antibody against TNFα is able to prevent allodynia induced by bortezomib but does not reduce neuropathy. Conclusion: Targeting TNFα might be useful in limiting patients' discomfort during bortezomib therapy.
Bortezomib is a drug with established anti-neoplastic activity against multiple myeloma and mantle cell lymphoma (1-4). Despite its efficacy, clinical use of bortezomib (particularly when delivered intravenously) is frequently limited by the onset of sensory peripheral neuropathy associated with neuropathic pain, while motor impairment is generally only sub-clinical (5, 6). Despite the fact that in most cases bortezomib-induced neuropathy is at least in part reversible (7) and that subcutaneous administration and less dose-intense administrations reduce its incidence, the capacity of bortezomib to damage the peripheral nervous system still remains of major concern since this might induce treatment withdrawal (8, 9).
The mechanisms for severe side-effect are still unclear. Bortezomib acts primarily by inhibiting protein degradation through binding to the 26S proteasome subunit (10). Downstream of proteasome inhibition, it also acts as a growth inhibitor by abrogating nuclear factor-KB (NF-KB) signaling eventually resulting in the production of tumor necrosis factor-α (TNFα) and bortezomib (BTZ) treatment induces activation of heat-shock proteins, generation of reactive oxygen species and induction of cell death pathways (11-15).
None of these events has ever been clearly associated with pathogenesis of the neuropathic pain induced by bortezomib administration in clinical practice. To study the onset of neurotoxicity of bortezomib and neuropathic pain, a reliable animal model, already extensively investigated and described, has been used (16-19).
It is accepted that neuropathic pain results from damage or inflammation of the nervous system, inducing painful sensory loss and hypersensitivity phenomena described as allodynia. TNFα is produced in response to diseases such as diabetes, tumorigenesis, inflammation, and tissue damage (20, 21). This cytokine has also been associated with the development of neuropathic pain (22-24), suggesting it could be a putative pharmacological target for treating this challenging medical condition. Several studies performed in rats and mice showed that through inactivation of TNFα, it is possible to reduce the enhanced sensitivity to mechanical and heat stimuli in models of painful neuropathy (25-27). For instance in inflammatory diseases, where the increased oxidant compounds levels are able to activate expression of TNFα gene and of other pro-inflammatory cytokines, the use of antibodies against TNFα has been useful in achieving an anti-inflammatory effect (21, 28).
On this basis, the present study was performed aiming to establish if the administration of a murine monoclonal antibody able to neutralize rat TNFα could prevent the development of neuropathic pain and peripheral neuropathy in rats induced by subchronic treatment with bortezomib.
Summary of the experimental plan. CNTO1081: Antibody against tumor necrosis factor-α.
Materials and Methods
Drugs and schedules of administration. Young adult female Wistar rats (175-200 g; Harlan Italy, Correzzana, Italy) were used. The care and husbandry of animals were in agreement with the Institutional guidelines in compliance with national (D.L. n. 116, Gazzetta Ufficiale della Repubblica Italiana, suppl. 40, Feb. 18, 1992 and subsequent modifications) and international laws and policies (EEC Council Directive 86/609, OJ L 358, 1, Dec. 12, 1987; Guide for the Care and Use of Laboratory Animals, U.S. National Research Council, 8th ed., 2011 available at http://grants.nih.gov/grants/olaw/Guide-for-the-care-and-use-of-laboratory-animals.pdf). The experiments were examined and approved by the Ethics Committee of the University of Milano-Bicocca (prot. n. 0023088 of November 8th, 2010) and by the Ethische Comissie Dierproeven at Janssen, Beerse, Belgium (Audit report from LAM Department/Ethics Committee of J&J of September 2nd, 2010). All the animals were housed in a limited-access animal facility at the University of Milano-Bicocca, where room temperature and humidity were set at 22±2°C and 55±10%, respectively. Artificial lighting provided 12 h light/12 h dark (7 a.m.-7 p.m.). A veterinarian with specific expertise in animal studies and the authority to withdraw the animal from the experiment examined any animal showing signs of distress.
Bortezomib (Janssen, Beerse, Belgium) solution was prepared before each administration; the powder was dissolved in a solution of 5% Tween 80, 5% ethanol 100% and 90% saline solution (a non-toxic vehicle according to the previously reported results) (29) and was administered intravenously at dose of 0.2 mg/kg three times a week for eight weeks. Murine monoclonal antibody specific for rat TNFα (CNTO1081) (Janssen R&D, Radnor, PA, USA) was dissolved in phosphate buffered saline (PBS) and was provided ready to use; anti-rat TNFα was administered intraperitoneally. In order to study the possible dose-related effect of anti-rat TNFα, two doses of the compound (2 mg/kg and 10 mg/kg) were tested in two separate experiments, as described in Table I. To ensure homogeneity among the treatment groups, baseline comparison was performed in nerve conduction velocity (NCV), dynamic test, plantar test and body weight assessment, with no evidence of any statistically significant difference.
Clinical signs and body weight. The general clinical condition of the animals was assessed daily during the treatment period, while the body weight was recorded twice weekly for toxicity assessment and adjustment of drug dose.
Neurophysiological assessment. At baseline and after four and eight weeks of treatment, the NCV was determined in the tail nerve of each animal as previously described (29, 30). Briefly, the animals were placed on a warmed mat and the NCV in the tail nerve was assessed by placing recording needle electrodes distally in the tail, while the stimulating needle electrodes were placed 5 cm and 10 cm proximally with respect to the recording point. The latencies of the potentials recorded at the two sites after nerve stimulation were determined (peak-to-peak) and the NCV was calculated accordingly. All the neurophysiological determinations were performed under standard conditions in a temperature-controlled room and under monitoring of animal vital signs and body temperature.
Histopathological examination. At the end of the treatment period (four days after the last administration) dorsal root ganglia (DRG) and sciatic nerves were collected and post-fixed in OsO4, epoxy resin embedded and used for light microscopy observation. For each animal, several semi-thin sections (1 μm) were prepared from randomly selected blocks and were stained with toluidine blue then examined with a Nikon Coolscope light microscope (Nikon Instruments, Calenzano, Italy).
Dynamic test. For each animal, mechanical tests were performed to determine the presence of allodynia. This test was performed before the beginning of the treatment (baseline) and after eight weeks of treatment using the dynamic plantar aesthensiometer test (model 37450; Ugo Basile Biological Instruments, Comerio, Italy). Rats were placed individually on an elevated wire mesh floor in a plexiglas chamber and allowed to acclimatize to the testing environment for at least 15 min. The tactile hind paw withdrawal threshold in rats was determined by calculating the mean value of six repeated applications of a 0.5 mm diameter metal filament to the plantar surface, which exerted a progressively increasing puncture pressure, reaching up to 50 g within 20 sec. Metal hairs were applied to the central region on the plantar surface avoiding the footpads and the pressure evoking a voluntary hind-paw withdrawal response (withdrawal latency) was recorded automatically. A cut-off time of 30 sec was used to prevent tissue damage in the absence of a response. Mechanical allodynia was defined as a significant decrease in the dynamic test threshold evoked by mechanical stimuli (16).
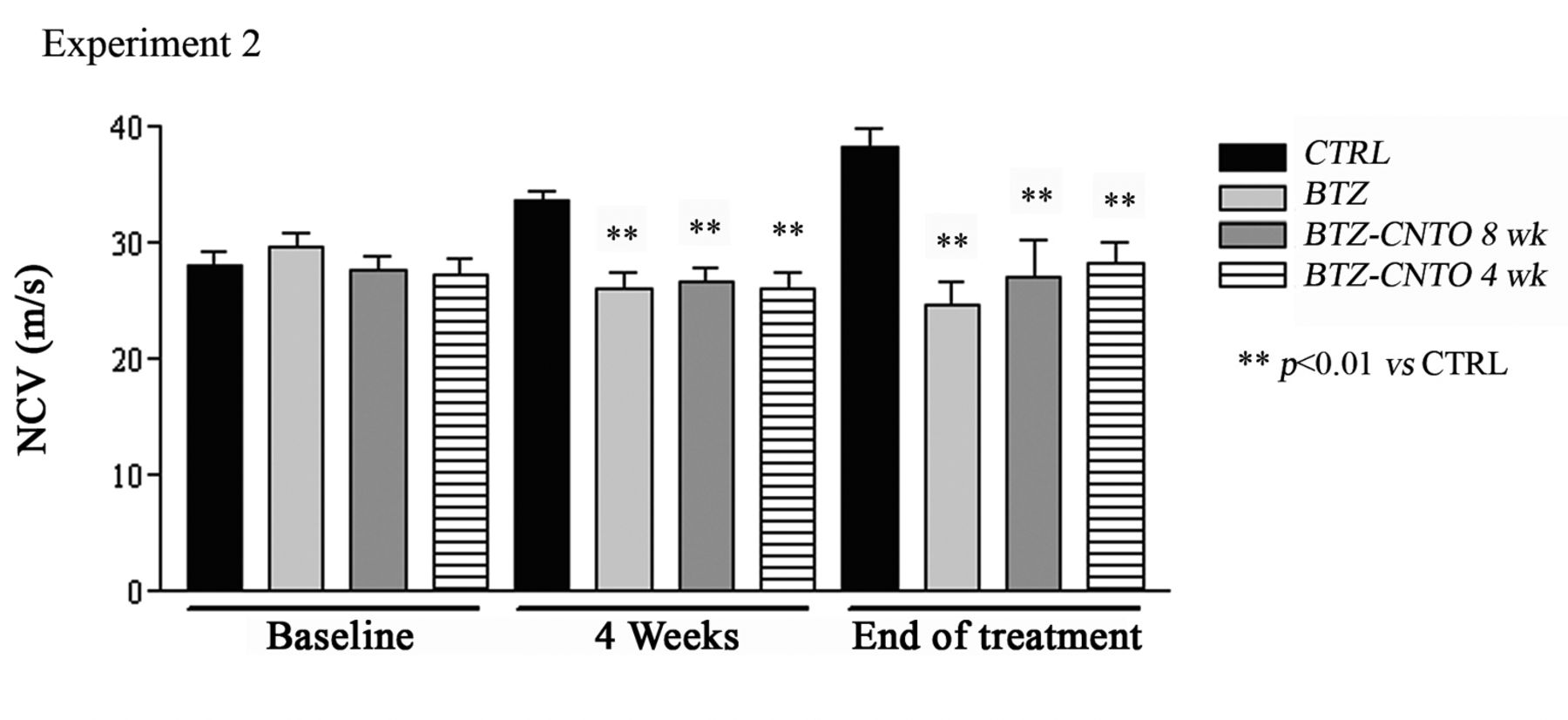
Changes in nerve conduction velocity (NCV). Measure of NCV at baseline and after four and eight weeks of treatment in rats treated with bortezomib (BTZ) alone and in co-treatment with anti-rat tumor necrosis factor-α (CNTO 1081) at a dose of 10 mg/kg (data are expressed as mean±standard deviation). See Table I for details of treatments.
Plantar test. Two hours after dynamic test evaluation, the response to noxious thermal stimulus was determined using a dynamic plantar analgesiometer (model 37370; Ugo Basile Biological Instruments), according to the procedure described by Hargreaves et al. (31) before starting the treatment and after eight weeks of treatment. During the plantar test the rats were placed into a plexiglas chamber for a 15-min acclimatization period followed by testing. A movable infrared radiant heat source (intensity of 40 W) was placed directly under the plantar surface on the hind paw and the time taken for hind paw withdrawal was monitored (withdrawal latency). The nociceptive threshold response to thermal stimulus was determined by calculating the mean value of four repeated measurements. A cut-off time of 30 sec was used to avoid any tissue damage (16).
Statistical evaluation. The differences in body weight, NCV, and dynamic and plantar tests were statistically compared with the results obtained for untreated animals using the one-way analysis of variance (ANOVA) and Tukey's multiple comparison test, as post-test (GraphPad Prism 3.0, GraphPad Software Inc., La Jolla, CA). The significance level was set at p<0.05.
Results
The experiments were carried out to assess if anti-rat TNFα administration was able to modify the severity of bortezomib-induced peripheral toxicity.
Mortality. In Experiment 1, three animals treated only with bortezomib died after 1, 21 and 23 administrations; two animals co-treated with bortezomib and anti-rat TNFα from the beginning (BTZ-CNTO 8 wk) died after 11 and 16 co-administrations with bortezomib, and four animals co-treated with anti-rat TNFα after four weeks of treatment with bortezomib (BTZ-CNTO 4 wk) died after 6, 7 and 8 co-administrations with bortezomib. In Experiment 2, four animals treated only with bortezomib died after 1, 17, 22 and 23 administrations, four in the BTZ-CNTO 8 wk group died after 1, 8, 13 and 18 co-administrations of bortezomib, and two co-treated in the BTZ-CNTO 4 wk group died after 4 and 5 co-administrations with bortezomib. Overall, these mortality rates are slightly higher than expected on the basis of the available data using exactly the same bortezomib model (16). Given the distribution of dead animals and the established tolerability to the antibody to TNFα (32, 33), mortality should reasonably be attributed to bortezomib administration.
Body weight changes. Weight changes in the treatment period were slightly different in the two experiments. In Experiment 1, after the first administration of bortezomib, a significant reduction of body weight (p<0.05) was observed in the BTZ and BTZ-CNTO 8 wk groups versus untreated control rats; at the end of the treatment, only BTZ-CNTO 8 wk and BTZ-CNTO 4 wk groups showed a significant reduction in body weight (p<0.05) versus controls. In Experiment 2, there was a less marked reduction of body weight in all these groups versus controls and only at the end of treatment did the animals in the BTZ group have a significant reduction in body weight (p<0.05) versus controls. However, in Experiment 2, animals in the BTZ group had a moderate weight loss in the last two weeks of treatment (mean weight loss 10 g), an event occasionally observed in previous studies (Meregalli, personal observation).
Neurophysiological evaluation. In both Experiment 1 and Experiment 2, the neurophysiological determinations evaluated at baseline showed that there was no statistically significant difference among groups. After four weeks and at the end of treatment, the same changes in NCV were found in both experiments, with a significant change in all bortezomib-treated groups versus controls. No effect of the co-administration of anti-rat TNFα was detected, even at the 10-mg/kg dose level (Figure 1).
Histopathological examination. The pathological evaluations were performed in both experiments on DRG and sciatic nerve obtained at the end of treatment from all groups of rats.
The morphological analysis by light microscopy of sciatic nerve confirmed our previous findings (16), showing mild axonopathy. In fact, only very rarely was mild de-generation of the myelin sheath observed in large myelinated fibers. Co-administration of anti-rat TNFα at any dose level had no effect on these pathological changes and did not induce any other remarkable effects on sciatic nerve fibers (data not shown).
Similarly, the analysis of DRG confirmed that bortezomib primarily induced morphological alteration in DRG satellite cells, with the appearance of vacuolization of the cytoplasm, while degenerating DRG neurons were also observed only very occasionally (16). As noted in the sciatic nerve, in the DRG the co-administration of anti-rat TNFα also had no effect on these pathological changes.
Behavioral tests. In both experiments, the determinations performed at baseline and at the end of treatment showed that there was no statistically significant difference among groups in the plantar test (data not shown).
In both experiments the determinations of dynamic test performed at baseline evidenced that there was not any statistically significant difference among groups (Figure 2). However in Experiment 1, at the end of treatment there was a marked reduction in latency, indicating the occurrence of mechanical allodynia in all bortezomib-treated groups versus untreated rats, even if this difference was not statistically significant (p=0.06, Figure 2a). By contrast, in Experiment 2, the reduction in latency observed at the end of treatment was statistically significant in bortezomib-treated versus control animals (p<0.05, Figure 2b). In the animals co-treated with CNTO1081 at the dose of 10 mg/kg from the beginning of bortezomib administration, there was a highly statistically significant protective effect of anti-rat TNFα, indicating reduced allodynia versus the group treated with bortezomib alone (p<0.01).
Discussion
Bortezomib is a cornerstone of the treatment of multiple myeloma and is also approved for the treatment of relapsed mantle cell lymphoma (1-4). Given its activity, the onset of severe side-effects, as in the case of painful sensory peripheral neuropathy (5, 6, 18), is a critical issue in clinical practice and investigations are required to deepen the knowledge of their pathogenesis and treatment. In fact, once established bortezomib-induced painful neuropathy is generally unresponsive to standard pain treatment and is perceived by the affected patients as severely limiting their daily activities and quality of life. A relevant step forward in overcoming this side-effect is represented by reduction in the incidence of bortezomib-induced peripheral neurotoxicity using the subcutaneous route, or through less frequent (weekly instead of twice-weekly) administration (9, 34-36). However, further investigation of the mechanisms underlying the capacity of bortezomib to damage the integrity of the peripheral nervous system is warranted.

Behavioral change after tumor necrosis factor-α (TNFα) treatment. Effect of treatment with bortezomib (BTZ) alone or in co-treatment with anti-rat TNFα at 2 mg/kg (a) and anti-rat TNFα at 10 mg/kg (b) on the withdrawal threshold response in the dynamic test (data are expressed as mean±standard deviation). See Table I for details of treatments.
Several clinical observations suggest that inflammation and immune-mediated events might be relevant actors in the onset and course of bortezomib-induced peripheral neuro-toxicity (37, 38). Moreover, analysis of single nucleotide polymorphisms in bortezomib-treated patients have identified pro-inflammatory genes (including TNFα-related genes) linked to the severity of bortezomib-induced peripheral neurotoxicity (39, 40), although the methodological approach to neurological assessment in these patients was not optimal according to recent revisions on the best methods to assess chemotherapy-induced peripheral neurotoxicity (41, 42), thus raising some possible concern as to the validity of the reported association (43, 44).
The present study was designed to test if, and to what extent, TNFα targeting using an inactivating monoclonal antibody could allow the severity of bortezomib-induced peripheral neurotoxicity to be modulated. In fact, not only is TNFα (as well as other pro-inflammatory cytokines) a major player in the onset and maintenance of neuropathic pain (22-24, 45), but a possible role of this cytokine has also recently been suggested in the pathogenesis of chemotherapy-induced peripheral neurotoxicity (46). A possible pathogenetic role of TNFα in nerve damage was also suggested in patients treated with hyperthermic isolated limb perfusion and TNFα for the treatment of advanced melanoma and sarcoma of the limbs showing mild, mainly sensory neuropathy in perfused limbs (47), but this has not been confirmed in other settings.
With this background, we tested the administration of two different doses of anti-rat TNFα given from the beginning of or after four weeks of bortezomib treatment, using an experimental paradigm able reliably to reproduce in rats the features of bortezomib-induced peripheral neurotoxicity in clinical practice. Using a multi-modal approach based on neurophysiological, behavioral and histopathological assessments, we have demonstrated that anti-rat TNFα administration has no evident effect on the onset and course of bortezomib-induced peripheral neuropathy, but has a remarkable and dose-dependent effect on mechanical allodynia. This result is potentially relevant, since it clearly indicates a role for TNFα in the course of bortezomib-induced neuropathic pain, while this cytokine apparently is not primarily involved in direct nerve and DRG damage induced by bortezomib. A possible explanation for this divergent effect might rely on a different site of action of TNFα in the course of bortezomib-induced peripheral neurotoxicity. For instance, since DRG neurons and satellite cells are clearly damaged by bortezomib administration (48), either a local or an indirect effect of TNFα on the spinal cord afferents targeting the dorsal horn neurons might be inferred and deserve to be further explored. Our results are in keeping with several studies which demonstrated high levels of TNFα expression in the standard model of chronic constriction injury of sciatic nerve in rats (49-51) and in nerve biopsies from patients with painful neuropathy (52). Moreover, several studies performed in rats and mice showed that inactivation of TNFα is able to reduce increased sensitivity to mechanical and heat stimuli induced in models of painful neuropathy (25-27).
The issue of TNFα as a possible target for therapeutic intervention in neuropathic pain is still unclear and at least in part could be dependent on the type of peripheral nerve damage. In literature, there are several clinical trials describing the opposite results with the use of commercial TNFα inhibitor (etanercept or infliximab) for the treatment of sciatica. Ohtori and colleagues showed that the administration of TNFα inhibitor is useful to reduce radicular pain caused by lumbar disc herniation and spinal stenosis in patients with low back and radicular leg pain (53). By contrast, Korhonen and colleagues performed two different clinical trails which tested the administration of infliximab in patients with acute/sub acute sciatica secondary to a herniated disc and observed that there was no significant reduction of lumbar radicular pain after such treatment (54, 55).
In conclusion, we have demonstrated that the co-administration of a neutralizing antibody against TNFα prevents allodynia, without reducing nerve or DRG damage. However, this beneficial effect suggests that a pharmacological-based approach might be useful in limiting pain-related discomfort in patients during bortezomib therapy; this needs to be carefully assessed in order to rule-out any possible interference with the anticancer activity of bortezomib if co-administered during chemotherapy (56, 57).
Footnotes
-
Conflicts of Interest
All authors at the University of Milan Bicocca have no conflict of interest instead J. Monbaliu and H. van de Velde are Janssen Research and Development employees.
- Received October 4, 2013.
- Revision received November 7, 2013.
- Accepted November 8, 2013.
- Copyright© 2013 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved





{kind=link}
{kind=link}