


Abstract
Background: The transcription factor SOX2 controls gene expression during development and has roles in both neurogenesis and gliogenesis. Materials and Methods: The current study utilized immunohistochemistry, immunofluorescence, molecular genetics (gene copy number analysis) and Western blotting to examine SOX2 expression in surgical samples from 133 brain gliomas of different grades of malignancy, and in cell lines from 16 glioblastomas. Results: Our results reveal a positive correlation between SOX2 expression and malignancy grade in gliomas and identify the hypercellular and hyperproliferative areas of glioblastomas as the areas with the highest SOX2 expression. SOX2 gene amplification was found in neurospheres and 14.4% and 11.1% of glioblastomas and anaplastic oligodendrogliomas, respectively. In contrast, SOX2 expression was not observed in neuronal tumors and varied in medulloblastomas depending upon their neuronal differentiation status. Conclusion: It is concluded that SOX2 is a marker for undifferentiated and proliferating cells and that its expression is up-regulated in the most anaplastic areas of glioblastomas and oligodendrogliomas. The hypothesis is that antigenic and genetic analogy exist between these areas and neurospheres and that glioblastoma stem cells may actually be dedifferentiated tumor cells that re-acquire a stem cell-like status.
Sex determining region Y-box 2 (SOX2) is a single-exon, intronless gene that is located on chromosome 3q26.3–q27 and encodes a 317 amino acid protein. SOX2 contains a high mobility group (HMG) DNA-binding domain and belongs to the SOX family of transcription factors (1). It is highly expressed during embryonic development but is down-regulated when neural cells exit the cell cycle and differentiate (2). SOX2 controls the expression of several important developmental genes (3) and plays a role in both neurogenesis and gliogenesis (4, 5), even though the mechanisms of neurogenesis inhibition are not fully understood (6). In chick spinal cord, the inhibition of SOX2 activity in neural progenitors leads to the inhibition of proliferation and to early neuronal differentiation (7). SOX proteins may sequester proneural proteins in complexes which lose the ability to bind to DNA or to trans-activate (8).
Early in development, SOX2 is present in neural tube cells and in neural stem cells (NSCs) (3). However, as development proceeds, the expression of SOX2 becomes restricted to the sub-ventricular zone (SVZ) and sub-granular zone (SGZ) of dentate gyrus. It is considered to be a marker of NSCs, because all SOX2 expressing cells also express nestin (9). During development, SOX2 expression is limited to proliferating cells, NSCs and progenitor cells. Upon differentiation, astrocytes lose SOX2 expression, but they preserve expression of genes in the SOXE sub-family (8). Upon becoming reactive astrocytes, they can switch the gene from quiescence to active. In neural progenitors, SOX2 inhibits neurogenesis and up-regulates the Notch signaling pathway (10). However, it can play a role in neurogenesis (11) and continues to be expressed in certain subpopulations of neurons, including neurons in the thalamus and striatum (5) and in and Purkinje cells (12).
In the adult brain, SOX2 expression is nearly undetectable (13), with the exceptions already mentioned (10). SOX2 is regulated via a number of mechanisms, including post-translational protein modification. For example, the acetylation of a lysine residue within the DNA-binding domain of SOX2 enhances its transport to the cytoplasm. In addition, SOX2 can be ubiquitinated and targeted for proteasomal degradation in embryonic cells in response to differentiation signals (14).
In nervous system tumors, SOX2 expression has been demonstrated in astroglial, oligodendroglial and ependymal, but not neuronal, lineages and has been shown to correlate inversely with differentiation (15, 16). It has been demonstrated in neurospheres from pediatric tumors (17-19) and, together with nestin, musashi-1 and CD133, in neurospheres from gliomas (20) and in gliomatosis cerebri (21); in gliomas, its expression increases with the grade of malignancy (22). Correspondingly, an overexpression of SOX2 mRNA has been recently demonstrated in most gliomas, with no expression in normal CNS (13). Silencing SOX2 in freshly derived glioblastoma tumor-initiating cells prevents their proliferation and inhibits tumorigenicity (23). On the other hand, SOX2 intervenes in the reversion of oligodendrocyte precursor cells to neural stem-like cells in culture and can be reactivated by breast cancer 1 (BRCA1) and Brahma protein (BRM1) through chromatin remodeling induced by platelet-derived growth factor (PDGF) (24).
In the present study, we wanted to clarify SOX2 expression, distribution and gene copy number status in normal nervous tissue, in a series of neuroepithelial tumors and in cell lines from primary glioblastomas (GBM) by immunohistochemistry, Western blotting and molecular biology.
Materials and Methods
Patients and tumor specimens. The study was carried out on: i) formalin-fixed and paraffin-embedded (FFPE) surgical samples from 171 brain tumors diagnosed according to the criteria of WHO (25) and listed in Table I; ii) five FFPE surgical samples of normal non-infiltrated peritumoral nervous tissue removed from around vascular malformations; iii) FFPE autopsy samples of the frontal, temporal, parietal, and occipital cortices, the basal ganglia and thalamus, the brain stem, the oblongata and the cerebellum, obtained from two patients who died after heart attacks.
All patients provided their informed consent and the study was carried out in accordance with approved ethical standards of the University Hospital of Turin, conforming to the provisions of the Helsinki Declaration.
Immunohistochemistry. Immunohistochemistry was carried out on the tumors listed in Table I, but in only 75 out of the 104 GBMs. Five-μm-thick sections were cut. Immunohistochemistry for a series of antibodies (Table II) was carried out using a Ventana Full BenchMark® automatic immunostainer (Ventana Medical Systems Inc., Tucson, AZ, USA) with the UltraView™ Universal DAB Detection Kit. Negative controls were performed by omission of the primary antibody. Heat-induced epitope retrieval (HIER) was performed with Tris-EDTA, pH 8.
Double staining for SOX2 (mouse monoclonal antibody) and glial fibrillary acidic protein (GFAP; rabbit polyclonal antibody) was carried out in peritumoral astrogliosis samples.
In vitro culture. In 16 out of the 75 glioblastomas, samples parallel to those for formalin fixation were washed, minced, and enzymatically dissociated (26), and primary cells were re-suspended both in Dulbecco's modified Eagle's medium (DMEM)/F-12 containing epidermal growth factor (EGF) and basic fibroblast growth factor (bFGF) and in DMEM with 10% fetal bovine serum (FBS) at 5% O2/CO2. Neurospheres (NS) developed in 6 cases, and adherent cells (AC) were observed in 9 cases. GBM reference cell lines were U87-MG and 010627 (from Galli R, DIBIT San Raffaele, Milan, Italy). NS and AC were transplanted into Nod SCID mice (Charles River, Calco, Italy) via the stereotactic injection of 2 μl of a 1×108/ml PBS cell suspension into the right striatum, according to Galli et al. (27). Clonogenic assays were carried out (28). After the addition of 3% FBS, NS differentiated into cells of astroglial, oligodendroglial and neuronal lineages.
Number, male: female ratio, mean age and range of all the samples.
Ethics statement. All animals were handled in strict accordance with good animal practice as defined by the relevant international (Directive 86/609/EEC and the recommendation 2007/526/EC from the European Community) and national (Legislative Decree 116/92 and law n. 413/1993) directives and according to protocol approved by the Animal Care and Use Committee of the H.S. Raffaele (IACUC #316) and communicated to the Ministry of Health and local authorities according to the Italian law. As a general rule, we applied all available methods to replace, reduce and refine animal use in our experimental plan. All experiments were terminated 140 days following treatment, or when significant sign of animal suffering became apparent. All interventions were performed under general anesthesia if suffering were expected. All animal experiments were performed in the central animal facility by trained and experienced personnel. Accreditation from the local authorities was obtained for all personnel involved in animal experiments.
Frozen sections. From the 16 GBMs used for cultures, cryostat sections were cut from samples frozen at –80°C.
Immunofluorescence. Immunofluorescence was performed for SOX2, nestin and GFAP on frozen sections, NS and AC. Goat anti-rabbit IgG conjugated to fluorescein isothiocyanate (FITC) and rabbit anti-mouse IgG conjugated to tetramethyl rhodamine isothiocyanate (TRITC) were used as secondary antibodies. Nuclei were counterstained with 4',6-diamidino-2-phenylindole (DAPI). Observation was carried out using a Zeiss Axioskop fluorescence Microscope (Zeiss, Oberkochen, Germany) equipped with an AxioCam5MR5c coupled to an Imaging system (AxioVision Release 4.5; Zeiss).
Western blotting analysis. Western blotting was performed on fresh frozen tissue from the selected 16 primary tumors, on NS and on AC. Proteins were extracted in RIPA lysis buffer containing protease and phosphatase inhibitor cocktails (Sigma-Aldrich, St. Louis, MO, USA), separated by sodium dodecyl sulphate - polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto polyvinylidene fluoride (PVDF) membranes (Immobilon-P; Millipore, Bedford, MA, USA). The antibodies used were against SOX2 (mouse monoclonal antibody) and nestin. A specific anti-α-tubulin antibody (LF-PA0146; LabFrontier, Seoul, Korea) was used to normalize sample loading and transfer. Proteins were detected by enhanced chemiluminescence (Millipore) and densitometric quantification of band intensities was performed using NIH Image J software (SB, NIMH, Bethesda, MD, USA).
Antibodies employed.
Molecular genetics. Genomic DNA was extracted from FFPE and frozen tumor samples using a standard phenol-chloroform protocol and from GBM cell lines using a QIAmp DNA Mini Kit (Qiagen GmbH, Hamburg, Germany). Quantitative real-time PCR for SOX2 was performed on a StepOne Real-Time PCR system using the StepOne Software v2.1 (Applied Biosystems, Foster City, CA, USA), according to the manufacturer's instructions. RnaseP was used as endogenous control. Multiplex PCRs were undertaken in a reaction volume of 20 μl containing 1X TaqMan® Genotyping Master Mix, 0.9 μM RnaseP primers, 0.2 μM RnaseP probe, 1 μl SOX2 TaqMan® Copy Number Assay (# Hs02719379_cn; Applied Biosystems) and 20 ng of genomic DNA. The target probe was FAM™ labeled and the endogenous control probe was VIC® labeled. Genomic DNA was amplified using the following PCR conditions: 95°C for 10 min, followed by 40 cycles of 95°C for 15 s and 60°C for 60 s. A no-template control was included in each assay and each sample was analyzed in triplicate. Quantification of SOX2 gene copy number was performed using the comparative ΔΔCt method and data analysis was carried out with CopyCaller software v1.0 (Applied Biosystems). Fold increase of the target SOX2 gene was calculated using 2ΔΔCt. Values of SOX2 gene copy number greater than 3 were defined as gene amplification, and values between 2 and 3 were considered to be gain of function.
Statistical analysis. Western blotting results were analyzed using the Student's t-test. Values of p<0.05 were considered statistically significant.
Results
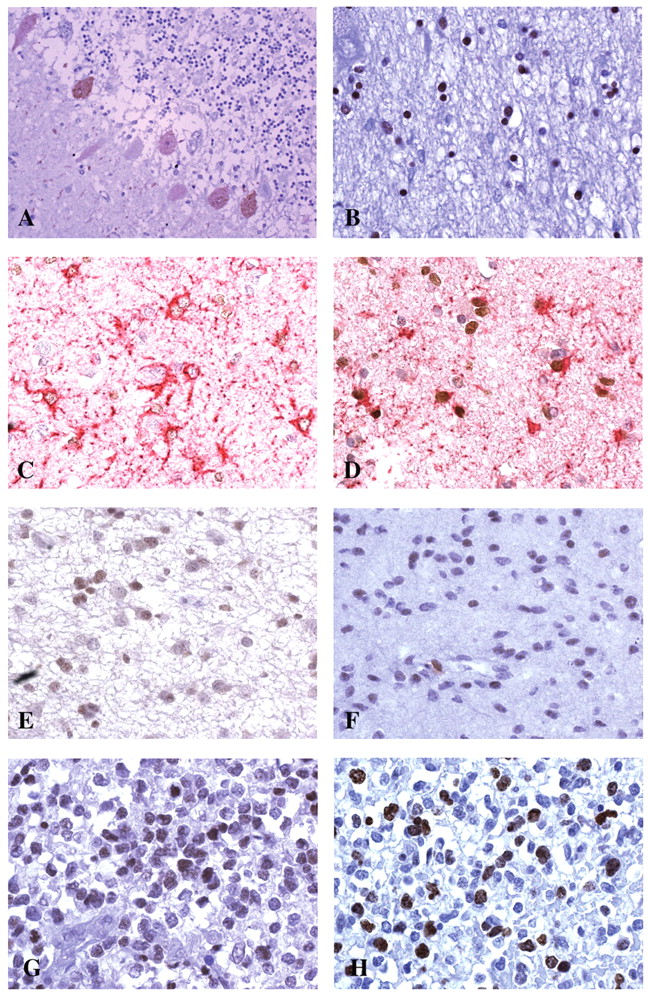
Immunohistochemistry. SOX2 was undetectable in almost all neurons in the normal brain, with the exception of Purkinje cells, which showed a weak nuclear and cytoplasmic staining (Figure 1A). Glia cells were also predominantly negative for SOX2 expression with the exception of some nuclei of scattered cells of thalamus, brain stem and hippocampus (Figure 1B). Peritumor astrogliosis was composed of two types of reactive astrocytes: i) large GFAP+ cells with long, thick processes and a clear large nucleus, and ii) smaller cells with short, thin processes. The larger astrocytes were more fibrillogenic and did not show mitosis; the smaller cells were less fibrillogenic and occasionally showed mitosis. The first type of reactive astrocytes, very likely longstanding, were found in nervous tissue surrounding vascular malformations; the second type were of recent development. Both astrocytic types were found in normal peritumoral tissue, as well as in normal tissue infiltrated by the tumor. The old fibrillogenic astrocytes were negative for SOX2 expression, but the younger astrocytes with short processes showed weak nuclear staining (Figure 1C, D). In diffuse astrocytomas and grade II oligodendrogliomas, we observed a low intensity staining (Figure 1E, F) in approximately 20-30% of nuclei. In anaplastic astrocytomas and oligodendrogliomas, in addition to this weak staining, there was a stronger staining pattern in up to 70% of nuclei (Figure 1G) and a variable proportion of intensely stained nuclei which corresponded to Ki-67/MIB.1-positive cells (Figure 1H). In GBMs, the percentage of intensely stained SOX2-positive nuclei was higher, between 50% and 100% (Figure 2A) and corresponded to a higher Ki-67/MIB.1 labeling index (LI). This picture was observed in a particular phenotype of GBM, characterized by high cell density with a high vessel density, including the external slope of circumscribed necroses, some invasion areas or perivascular cuffings (Figure 2B). In these areas, high SOX2 expression and high Ki-67/MIB.1 LI were associated with a higher nestin expression than GFAP (Figure 2C), and than the remainder of the tumor, especially around vessels.
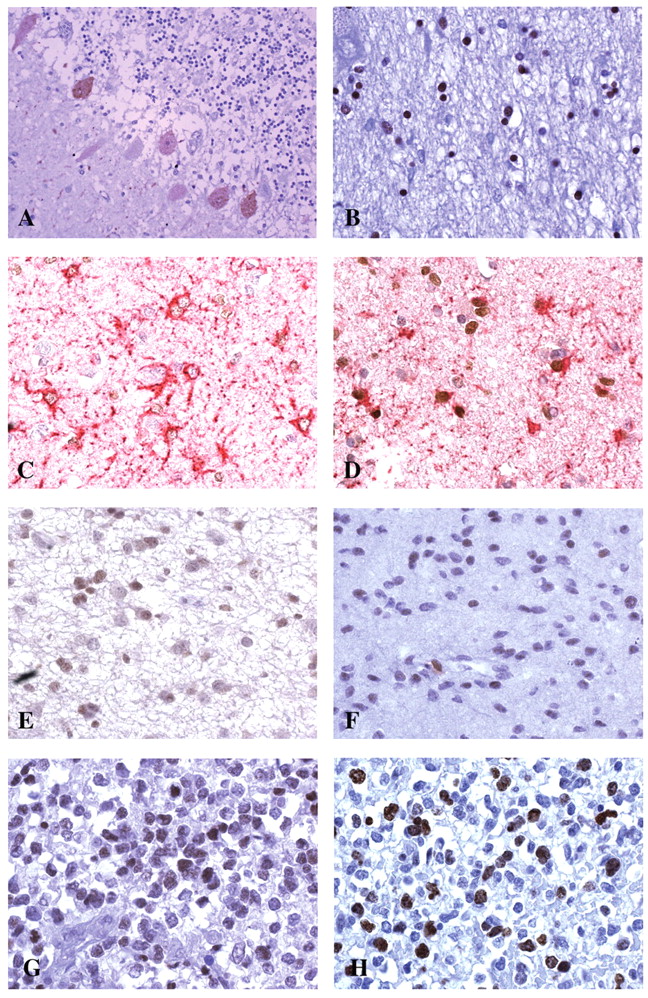
Immunohistochemistry. A: Positive SOX2-staining in Purkinje cells. DAB, ×200. B: Positive glial nuclei in thalamus. DAB, ×400. C: Long-lasting fibrous reactive astrocytes with SOX2-negative nuclei. Double staining GFAP-SOX2, ×400. D: Young reactive astrocytes with short processes. SOX2-positive nuclei. Double staining GFAP-SOX2, ×400. E: Diffuse astrocytoma. Few positive nuclei. DAB, ×400. F: Oligodendroglioma. Scattered positive nuclei. DAB, ×400. G: Anaplastic oligodendroglioma. Many SOX2-positive nuclei. DAB, ×400. H: Section from the same area of G with many Ki-67/MIB.1-positive nuclei. DAB, ×400.

Immunohistochemistry and immunofluorescence. A: Intensely SOX2-positive nuclei in hyper-cellular area of glioblastoma. DAB, ×400. B: Perivascular cuffing of intensely positive nuclei cells. DAB, ×200. C: Intensely nestin-positive perivascular cell cuffing. D: Medulloblastoma-positive cells around a negative pale island. DAB, ×200. E: Central neurocytoma. Negative staining. ×400. F: Meningioma, area with positive nuclei. DAB, ×400. G: Neurospheres from a glioblastoma. Intense nuclear green fluorescence. Immunofluorescence, ×400.
In medulloblastomas, SOX2 staining was either positive or negative (Figure 2D), according to the occurrence of areas with neuronal differentiation, like pale islands, demonstrated by synaptophysin expression. The only neurocytoma was negative for SOX2 (Figure 2E). A variable staining was observed in ependymomas, with some high intensity areas (data not shown), and a variable and low intensity staining was found in nuclei of meningiomas (Figure 2F). Immunohistochemistry results for the glioma series are summarized in Table III.
Tumors developed in mice after transplantation of cell lines showed similar patterns of SOX2 nuclear staining to that of the primary tumors.
Immunohistochemistry semiquantitative intensity evaluation and mean percentages in the glioma series.

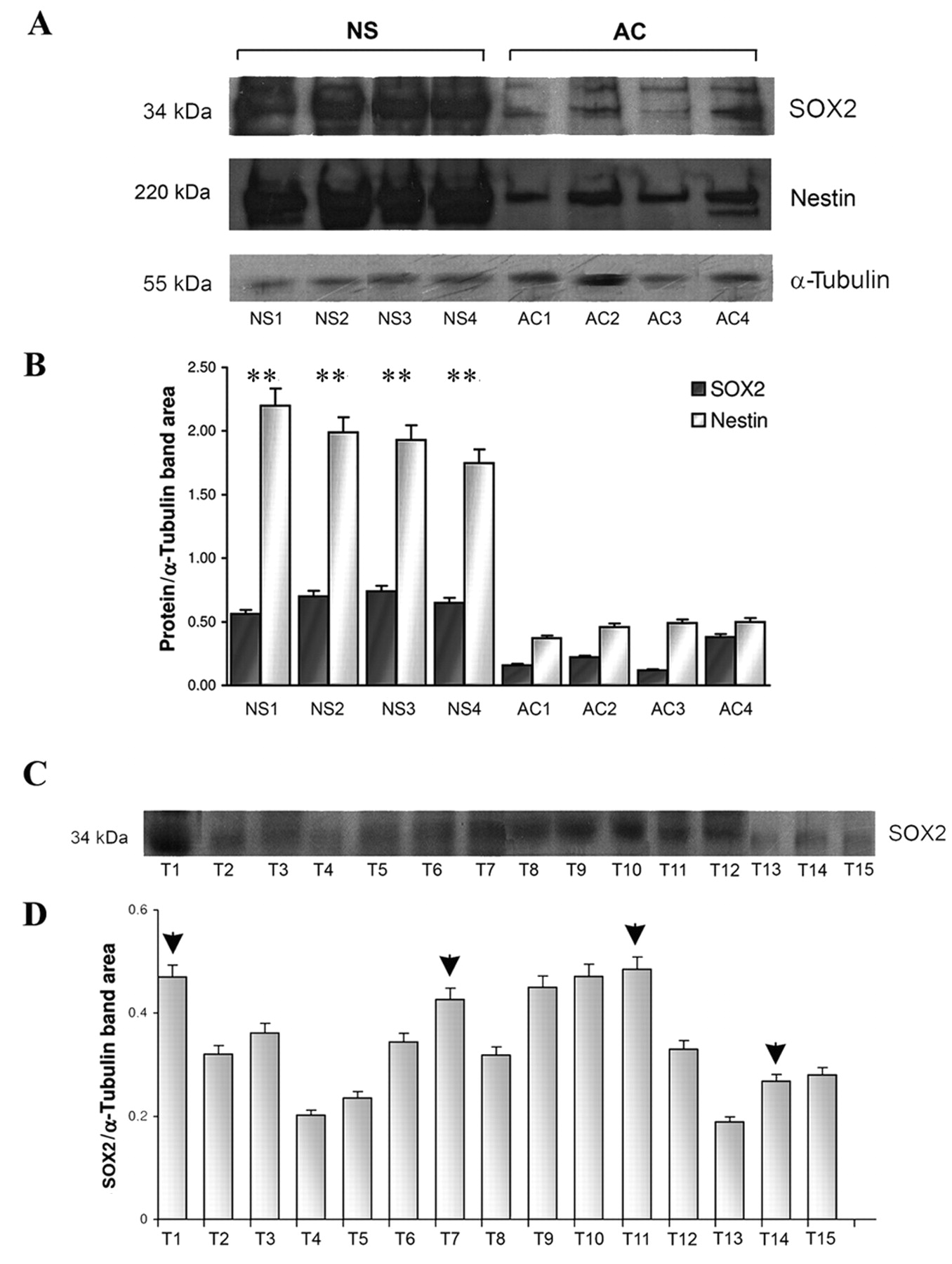
Representative Western blot of SOX2 and nestin. A: Higher protein expression in neurospheres (NS) than in adherent cells (AC), both for SOX2 and nestin; B: densitometric quantification, **p<0.01. C: SOX2 protein expression in GBM primary tumors; D: densitometric quantification. Arrow heads indicate primary tumors from which NS were generated.
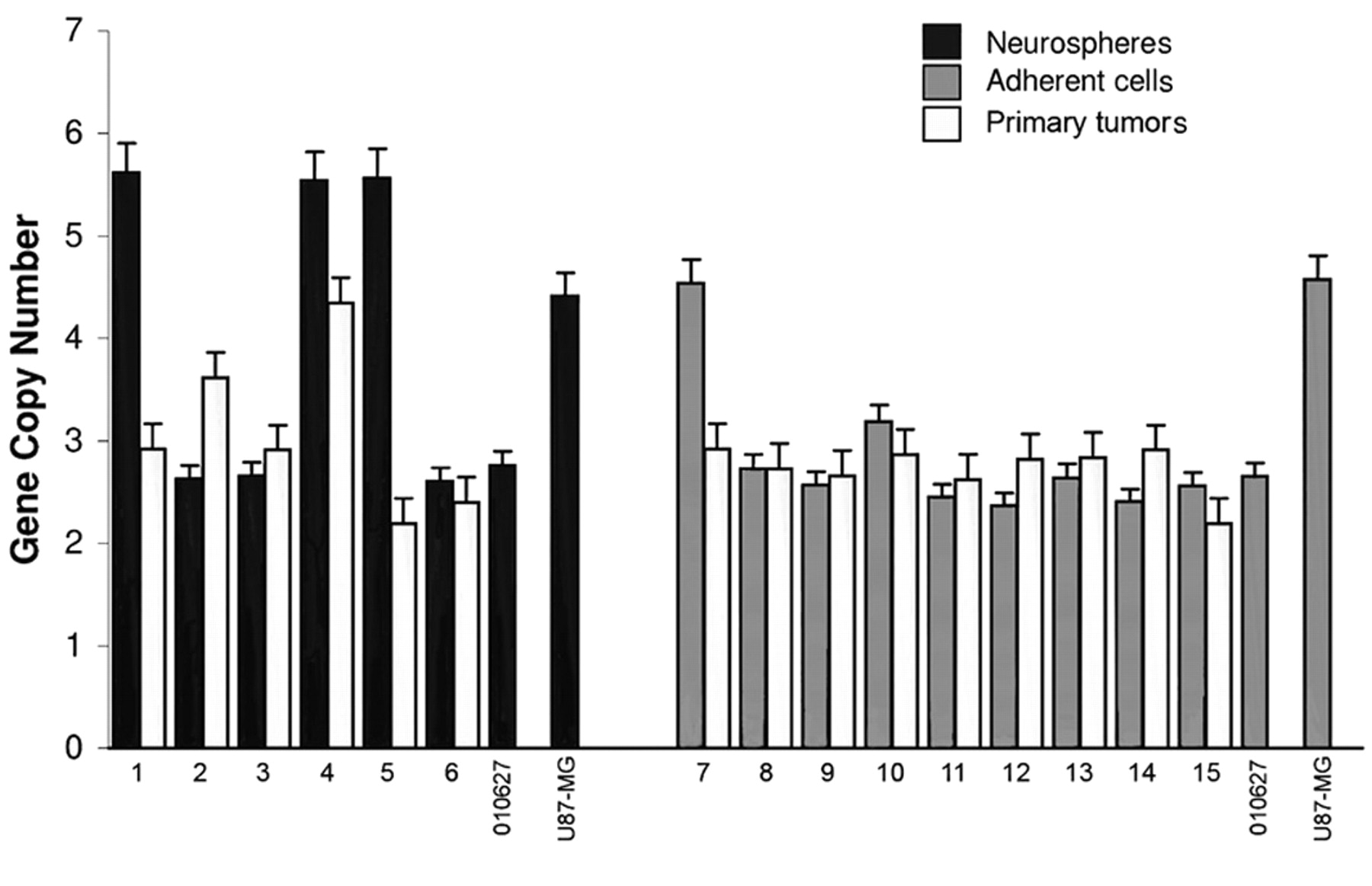
TaqMan® gene copy number analysis. Results for SOX2 TaqMan® Copy Number Assay in the whole glioma series.
Amplification percentages in the glioma series.
Immunofluorescence. Results of immunofluorescent staining varied according to the antibody used. With the rabbit antibody, cytoplasmic staining was observed for SOX2. In contrast, the mouse antibody revealed a staining distribution more similar to that observed by immunohistochemistry. In cell lines, with mouse antibody there was a constant nuclear staining of high intensity in NS (Figure 2G), whereas in AC, the nuclei were irregularly and weakly stained and an irregular staining of the cytoplasms was also observed.
Western blotting analysis. Western blotting analysis for SOX2 revealed a single band at approximately 34 kDa (Figure 3A, C). Quantitative evaluation demonstrated a remarkably higher SOX2 expression in primary tumors from which NS developed and in NS in comparison with AC (Figure 3B, D). Nestin showed a parallel pattern of distribution.
Molecular genetics. TaqMan® gene copy number analysis revealed amplification of SOX2 gene in 3 out of the 6 NS, in 2 out of the 9 AC and in 2 out of the 6 primary tumors generating NS. The remaining NS, AC and primary tumors from which cell lines developed showed a gain of function. Both NS and AC from 010627 cell line revealed a gain of function, whereas NS and AC from U87-MG cell line resulted in amplified expression (Figure 4). In the series of gliomas, 15 out of the 104 GBMs (14.4%) and 2 out of 18 anaplastic oligodendrogliomas (11.1%) showed SOX2 amplification. Diffuse and anaplastic astrocytomas as well as oligodendrogliomas showed no gene amplification (Table IV).
Discussion
Consistent with other reports from the literature, our data demonstrate that SOX2 is expressed in the nucleus of undifferentiated and proliferating cells. We did not observe cytoplasmic location of SOX2, except in Purkinje cells and AC, and when using the rabbit antibody in immunofluorescence studies. Purkinje cells belong to a group of neurons, such as those of cortex, thalamus and striatum, in which SOX2 is expressed during adulthood (4, 5, 12). Why this may happen is not exactly known. Previous studies, aimed at identifying the functions of SOX2 in adult brain, including its relationship with anophthalmia and seizures (4, 5), did not reach clear-cut conclusions, other than the fact that SOX2 has an unexpected role in neuronal function and maintenance (11, 29).
There is one notable exception to the nuclear localization of SOX2 in early embryogenesis, when SOX2 is shuttled from the cytoplasm to the nucleus at the two-cell stage, at which point it remains cytoplasmic only in trophoectodermic cells (3). It has been demonstrated that the nuclear or cytoplasmic localization of SOX2 may depend on the antibody used, at least in adipose stem cells (30). We did not investigate this question in our material. In the case of AC, the cytoplasmic localization of SOX2 has been interpreted to result from its differentiation-induced proteolytic degradation (14). On the other hand, nuclear SOX2 expression is also lost upon differentiation of AC (2). It is possible that this explains the weaker SOX2 expression observed by Western blotting in AC when compared to NS.
In the chick spinal cord, the inhibition of SOX2 activity induces the exit of neural progenitors from the cell cycle and results in their differentiation (6, 7, 31). In summary, SOX2 inhibits neurogenesis, but not gliogenesis and is expressed in NS but disappears upon differentiation (10). These results are consistent with our findings of negligible SOX2 expression in neurons and glial cells in the adult brain. It is difficult to explain the weak SOX2-positive signal observed in some glia cells in thalamus, hippocampus and brain stem, unless these cells are not yet fully differentiated. In addition to SOX2 staining being observed in some adult glial cells, Bergmann's glia have also been reported to be positive for SOX2 (32). There is still some controversy regarding the identity of SOX2-positive astrocytes within the adult brain. For example, some have argued that SOX2-positive astrocytes observed in the adult rat brain are `neurogenic' astrocytes residing within a `neurogenic' niche (33). Interestingly, it has been shown in culture that rat astrocytes can resume proliferation after injury (10). This is in line with our observation of increased SOX2 expression in early, still proliferating astrocytes in comparison with quiescent, fibrillogenic ones.
In brain tumors, SOX2 expression is positively correlated with the malignancy grade (13, 15, 16, 22). Our results are consistent with this correlation and confirm the lack of SOX2 expression in neuronal tumors. In fact, we did not observe SOX2 expression in neurocytoma or in neuronally differentiated medulloblastomas. Of particular interest is the intense SOX2 nuclear staining observed in hypercellular areas with high vessel density, including the external slope of circumscribed necroses and invasion areas of GBMs. In these areas, we observed a correlation between SOX2 expression, Ki-67/MIB.1 LI and nestin expression. We have previously hypothesized that the opposing behavior of nestin and GFAP in these areas is due to dedifferentiation with embryonic regression (34). These very areas have been hypothesized to contain brain tumor stem cells or to give origin to cells exhibiting many of the features of stem cells, i.e. in a stem cell-like status (35). This observation may explain the amplification of SOX2 in NS, the SOX2 gene amplification of certain GBMs, and the more intense SOX2 expression by Western blotting observed in comparison with other gliomas (data not shown). Previous work has shown that SOX2 amplification is observed in 11.8% of cases of prostate cancer (36) and in a certain percentage of lung and esophageal squamous carcinomas, depending upon the pluripotency of cells (37-40). Areas of anaplastic oligodendrogliomas overexpressing SOX2 and with a high Ki-67/MIB.1 LI can have an analogous interpretation.
The aforementioned hypercellular areas in GBMs may correspond to the areas of highest tumor cell dedifferentiation, which occurs as a consequence of accumulation of mutations or of epigenetic events, and contain new malignant clones arising through anaplasia. These very areas are characterized by the expression of antigens of stemness, such as nestin, CD133, musashi-1 (20, 22, 41) and they have been shown to occur more frequently in tumors forming NS in culture than in those giving origin to AC only (35). In this regard, it can be emphasized that SOXB1 proteins are essential for the activation of the nestin enhancer and that nestin and SOX2 are functionally linked (6), and that both are overexpressed in the aforementioned areas. It is possible that SOX2 is sufficient to induce pluripotency and oncogenic activity (37) or that it plays a permissive role (42). Our findings are in line with those of Ma et al. (22) and follow our previous observations on GBM cell lines from the same tumors (35).
SOX2 is expressed in gliomas in relation to the grade of malignancy. In glioblastoma it is intensely expressed in the most malignant component of the tumor and in highly proliferating cells of oligodendroglioma. In a certain percentage of glioblastomas and oligodendrogliomas it is amplified. Generally, in malignant gliomas its intense positive staining distributes as Ki-67/MIB.1-positive nuclei. In areas with an intense staining, SOX2 is mostly amplified. In cultures it is amplified in neurospheres. The hypothesis that a genetic and antigenic analogy exists between neurospheres and the most anaplastic areas of gliomas is confirmed by SOX2 findings.
Acknowledgments
This study was supported by a grant from Compagnia di San Paolo, Turin, Italy.
- Received November 24, 2011.
- Revision received March 11, 2011.
- Accepted March 15, 2011.
- Copyright© 2011 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Canonical BMP2/4 signaling degrades SOX2 in glioblastoma propagating cells
- Dichaete, a Sox2 homologue, prevents activation of cell death in multiple developmental contexts
- Reprogramming Transcription Factors Oct4 and Sox2 Induce a BRD-Dependent Immunosuppressive Transcriptome in GBM-Propagating Cells
- A Sox2:miR-486-5p Axis Regulates Survival of GBM Cells by Inhibiting Tumor Suppressor Networks
- Single-Cell Transcriptome Analysis of Lineage Diversity and Microenvironment in High-Grade Glioma
- Transcriptional repressor GATA binding 1-mediated repression of SRY-box 2 expression suppresses cancer stem cell functions and tumor initiation
- Activation of SOX2 Expression by BRD4-NUT Oncogenic Fusion Drives Neoplastic Transformation in NUT Midline Carcinoma
- Non-invasive neural stem cells become invasive in vitro by combined FGF2 and BMP4 signaling
- Sox2 protects neural stem cells from apoptosis via up-regulating survivin expression