


Abstract
We recently showed that poly(ADP-ribose) polymerase (PARP) inhibitors exert their cytotoxicity primarily by trapping PARP-DNA complexes in addition to their NAD+-competitive catalytic inhibitory mechanism. PARP trapping is drug-specific, with olaparib exhibiting a greater ability than veliparib, whereas both compounds are potent catalytic PARP inhibitors. Here, we evaluated the combination of olaparib or veliparib with therapeutically relevant DNA-targeted drugs, including the topoisomerase I inhibitor camptothecin, the alkylating agent temozolomide, the cross-linking agent cisplatin, and the topoisomerase II inhibitor etoposide at the cellular and molecular levels. We determined PARP-DNA trapping and catalytic PARP inhibition in genetically modified chicken lymphoma DT40, human prostate DU145, and glioblastoma SF295 cancer cells. For camptothecin, both PARP inhibitors showed highly synergistic effects due to catalytic PARP inhibition, indicating the value of combining either veliparib or olaparib with topoisomerase I inhibitors. On the other hand, for temozolomide, PARP trapping was critical in addition to catalytic inhibition, consistent with the fact that olaparib was more effective than veliparib in combination with temozolomide. For cisplatin and etoposide, olaparib only showed no or a weak combination effect, which is consistent with the lack of involvement of PARP in the repair of cisplatin- and etoposide-induced lesions. Hence, we conclude that catalytic PARP inhibitors are highly effective in combination with camptothecins, whereas PARP inhibitors capable of PARP trapping are more effective with temozolomide. Our study provides insights in combination treatment rationales for different PARP inhibitors.
Introduction
Since the discovery of the synthetic lethality of poly(ADP-ribose) polymerase (PARP) inhibitors in BRCA-deficient cells (Bryant et al., 2005; Farmer et al., 2005; McCabe et al., 2006; Helleday, 2011; Lord and Ashworth, 2012), the mechanism by which PARP inhibitors exert their cytotoxicity has been dominantly interpreted by an accumulation of unrepaired single-strand breaks (SSBs) resulting from catalytic PARP inhibition. This interpretation has recently been revisited after the demonstration that PARP inhibitors also trap PARP1- and PARP2-DNA complexes at DNA damage sites that arise spontaneously and/or are produced by the classic alkylating agent, methyl methanesulfonate (MMS) (Murai et al., 2012b). The fact that PARP1-depleted cells become tolerant to PARP inhibitors also supports the cytotoxic mechanisms of PARP trapping (Liu et al., 2009; Pettitt et al., 2013). PARP trapping is not merely interpreted as resulting from catalytic PARP inhibition, which prevents dissociation of PARP from DNA and is required for repair completion (Satoh and Lindahl, 1992). Indeed, BMN 673 (see Murai et al., 2014), olaparib (AZD-2281), and niraparib (MK-4827) are much more effective than veliparib (ABT-888) for PARP trapping at concentrations where BMN 673, olaparib, niraparib, and veliparib fully inhibit PARylation (Murai et al., 2012b, 2014). Based on the fact that olaparib and niraparib are much more cytotoxic than veliparib as single agents, it is plausible that PARP trapping is more cytotoxic than unrepaired SSBs caused by the absence of PARylation (Murai et al., 2012b, 2014). Chemical differences in drug structures may cause different allosteric effects between the PARP catalytic and DNA-binding domains, and we have proposed to classify PARP inhibitors based on their dual molecular mechanisms of action: catalytic inhibition and trapping of PARP (Murai et al., 2012b, 2014; Fojo and Bates, 2013).
Combinations of different PARP inhibitors with a broad spectrum of genotoxic drugs are in clinical trials. These combinations include alkylating agents (temozolomide), topoisomerase I inhibitors (the camptothecin derivatives topotecan and irinotecan), topoisomerase II inhibitors (etoposide), and cross-linking agents (cisplatin) (Rouleau et al., 2010; Kummar et al., 2012; Curtin and Szabo, 2013). However, based on the fact that not all PARP inhibitors act similarly (Murai et al., 2012b, 2014; Fojo and Bates, 2013), it is critical to rationalize the most relevant combinations by choosing which PARP inhibitor and which chemotherapeutic agent act most effectively. It is also important to elucidate which combinations induce PARP trapping. Under such circumstances, highly potent PARP-trapping drugs should be more effective than simple catalytic PARP inhibitors (olaparib > veliparib). On the other hand, if the synergistic effect is caused by catalytic PARP inhibition, veliparib should be comparable to olaparib.
In this study, we compared olaparib and veliparib in combination with four drugs from different therapeutically relevant classes (temozolomide, camptothecin, cisplatin, and etoposide) to evaluate the potential and rationale for each combination. To determine whether potentiation was related to PARP catalytic inhibition or trapping, we used genetically modified chicken lymphoma DT40 cells (Buerstedde and Takeda, 1991; Maede et al., 2014), as well as human cancer cell lines, and measured olaparib- and veliparib-induced PARP-DNA complexes (PARP trapping). We chose human prostate cancer cells (DU145) and human glioblastoma cells (SF295) from the NCI60 cell line panel because, in our previous studies, these cell lines showed differential responses to veliparib and olaparib with respect to drug sensitivity and PARP trapping (Murai et al., 2012b, 2014).
Materials and Methods
Cell Lines and Drugs.
DT40 cell lines were obtained from the Laboratory of Radiation Genetics Graduate School of Medicine at Kyoto University (Kyoto, Japan). Human prostate cancer cells (DU145; sex: male) and human glioblastoma cells (SF295; sex: female) were obtained from the National Cancer Institute Developmental Therapeutics Program (Frederick, MD). Olaparib, veliparib, and camptothecin were obtained from the Drug Synthesis and Chemistry Branch, Developmental Therapeutics Program, and Division of Cancer Treatment and Diagnosis, National Cancer Institute (Bethesda, MD). Temozolomide (T2577) and cisplatin (P4394) were purchased from Sigma-Aldrich (St. Louis, MO). Drug stock solutions were made in dimethylsulfoxide at 10 mM for olaparib and veliparib, 10 μM for camptothecin, 100 mM for temozolomide and in 0.75 M NaCl (pH 5) at 5 mM for cisplatin. Drug stock solutions were stored at −20°C in the dark and diluted in culture medium immediately before use. MMS (10%) was prepared fresh from 99% MMS (129925; Sigma-Aldrich) in phosphate-buffered saline, and diluted in culture medium immediately.
Immunoblotting.
Ten million DT40 cells with 10 ml of medium in a 15-ml tube or semiconfluent human cells with 5 ml of medium in a six-well dish were treated as indicated for 30 minutes or 4 hours, respectively, and then cells were collected. To prepare subcellular chromatin fractions, we used a Subcellular Protein Fractionation Kit from Thermo Fisher Scientific (Rockford, IL) following the manufacturer’s instructions. Immunoblotting was carried out using standard procedures (Murai et al., 2012b). Densitometric analyses of immunoblots were carried out using ImageJ software (National Institutes of Health). Each band blotted against PARP1 or histone H3 was selected with the rectangular selection tool, measured the intensity (area × mean), and the intensity of background was subtracted from the intensity of each band. The intensity of the PARP1 band was divided by the intensity of the corresponding histone H3.
Antibodies.
Rabbit polyclonal anti-PARP1 antibody (sc-7150) and mouse monoclonal anti–proliferating cell nuclear antigen (anti–PCNA) antibody (sc-56) were from Santa Cruz Biotechnology (Santa Cruz, CA). Rabbit polyclonal anti–histone H3 antibody (07-690) was from Upstate Biotechnology (Lake Placid, NY). Rabbit polyclonal anti–poly(ADP-ribose) (PAR) polymer antibody (336-BPC-100) was from Trevigen (Gaithersburg, MD). Secondary antibodies were horseradish peroxidase–conjugated antibodies to rabbit or mouse IgG (GE Healthcare, Chalfont St. Giles, Buckinghamshire, UK).
Immunoassay for PAR Substrate.
The validated chemiluminescent immunoassay for PAR using commercially available reagents was performed (for detailed laboratory procedures, go to http://dctd.cancer.gov/ResearchResources/biomarkers/PolyAdenosylRibose.htm).
Measurement of Cellular Sensitivity to Drug Treatment.
Cells were continuously exposed in triplicate to the indicated drug concentrations for 72 hours. For DT40 cells, 200 cells were seeded into 384-well white plates (PerkinElmer Life and Analytical Sciences, Waltham, MA) in 40 μl of medium per well (Murai et al., 2012a; Maede et al., 2014). For human cells, 1500 DU145 cells or 500 SF295 cells were seeded in 96-well white plates (PerkinElmer Life and Analytical Sciences) in 100 μl of medium per well. Cell viability was determined using the ATPlite 1-step kit (PerkinElmer Life and Analytical Sciences). In brief, 20 or 50 μl of ATPlite solution for 384-well or 96-well plates, respectively, was added to each well. After 5 minutes, luminescence was measured by EnVision 2104 Multilabel Reader (PerkinElmer Life and Analytical Sciences). The ATP concentration in untreated cells was defined as 100%. Viability (percentage) of treated cells was defined as (ATP concentration in treated cells/ATP concentration in untreated cells) × 100.
Flow Cytometric Analysis of Cell Cycle Progression.
Cells were pulse labeled with 50 μM bromodeoxyuridine during the last 20 minutes of drug treatment. Cells were harvested, fixed in 70% ethanol, and stored at −20C° for 10 minutes. Cells were incubated for 30 minutes at room temperature in 2 N HCl–0.5% Triton X-100 to allow DNA denaturation. The cells were washed twice with phosphate-buffered saline containing 0.5% Tween 20 and 0.5% bovine serum albumin. Cells were incubated for 1 hour at room temperature with a fluorescein isothiocyanate–conjugated anti-bromodeoxyuridine antibody (BD Biosciences, Franklin Lakes, NJ) and treated with 0.5 mg/ml RNase A and 5 μg/ml propidium idodide. Samples were analyzed with a flow cytometer (FACScan; BD Biosciences) using the CellQuest software (BD Biosciences).
Analysis of Combination Effects.
The synergism analysis for the combination effects was conducted using the Chou-Talalay method (Chou, 2010). The combination index (CI) of each combination treatment was calculated using CalcuSyn software (Biosoft, Inc., Cambridge, UK), and CI 0.3–0.7, CI 0.1–0.3, and CI < 0.1 were defined as synergism, strong synergism, and very strong synergism, respectively (Chou, 2006).
Statistical Analyses.
Results are presented as means ± S.D. (n ≥ 3) or as means (n = 2). Differences between samples were assessed using the unpaired t test. All analyses were conducted with Prism 5.0 (GraphPad Software, La Jolla, CA). P values were two-sided and considered statistically significant when P < 0.05.
Results
Olaparib Potentiates Temozolomide Better than Veliparib.
Temozolomide is a commonly used anticancer drug, which, similar to MMS, induces base damage resulting in abasic sites and single-strand breaks. Synergy between temozolomide and PARP inhibitors has been reported by independent studies (Boulton et al., 1995; Horton and Wilson, 2013; Kedar et al., 2012). To compare olaparib and veliparib, we exposed wild-type and PARP1-deficient (PARP1−/−) DT40 cells to temozolomide with or without the PARP inhibitors (Fig. 1, A and B). Note that PARP1−/− DT40 cells are equivalent to PARP1 and PARP2 dual-knockout cells because of genetic lack of PARP2 in avian cells (Hochegger et al., 2006; Murai et al., 2012b). Consistent with the widely accepted concept that PARP1 is necessary for SSB repair, we found that PARP1−/− cells were hypersensitive to temozolomide (compare upper and lower panels of either Fig. 1, A or B). The addition of 10 μM olaparib or veliparib had no impact on PARP1−/− cells, confirming a lack of off-target effect of either drug (Murai et al., 2012b). Olaparib synergistically sensitized wild-type cells in a concentration-dependent manner, with a low combination index (CI < 0.3; Supplemental Table 1) at several points, and the sensitization with 0.1 μM olaparib went beyond that of PARP1−/− cells (compare the panels of Fig. 1A). Veliparib also synergized the cytotoxicity of temozolomide with a low CI (<0.3; Supplemental Table 1), but at a higher concentration than olaparib. Moreover, a high concentration of veliparib (10 μM; >100-fold higher than olaparib) was required to go beyond the sensitivity of PARP1−/− cells.
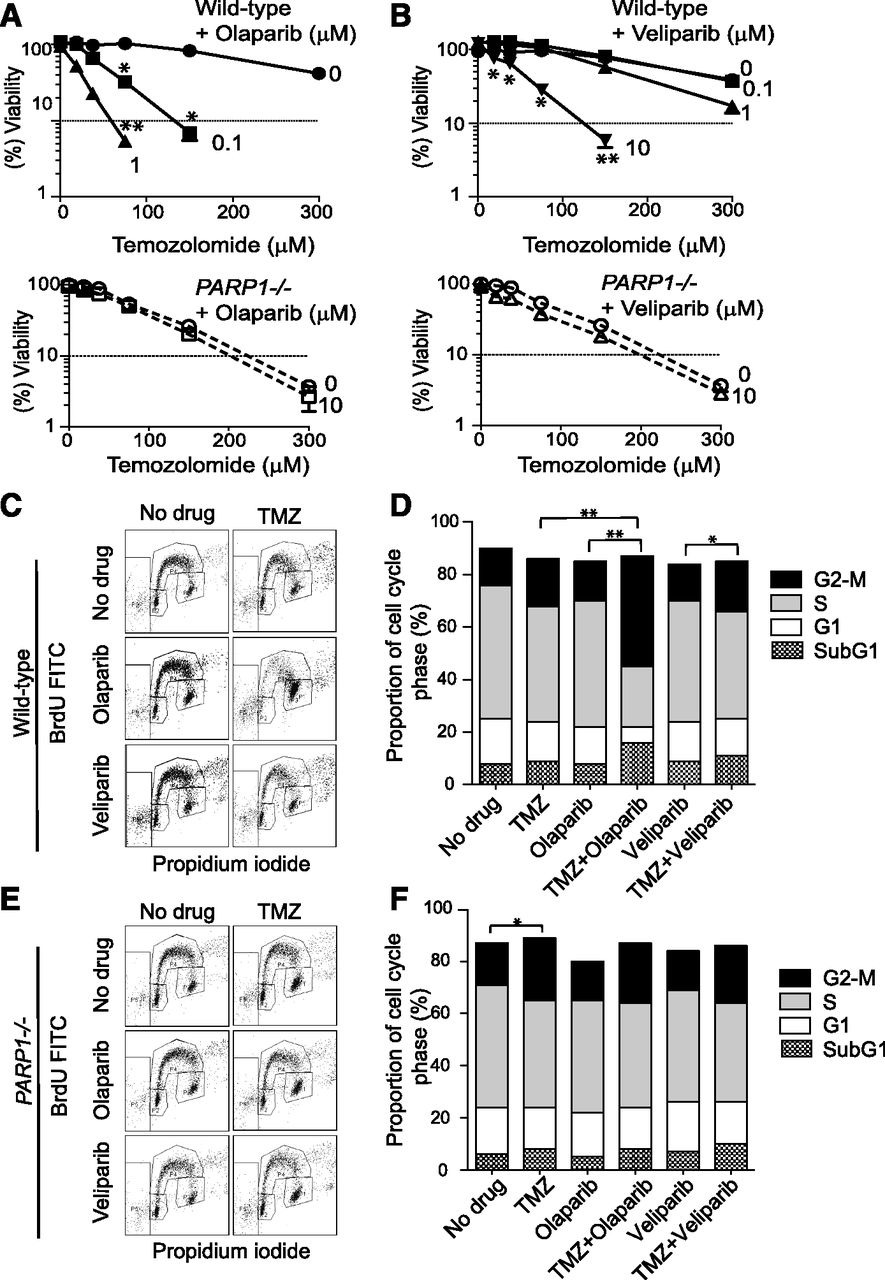
Olaparib augments the cytotoxicity of temozolomide better than veliparib and PARP deficiency. (A and B) Viability curves of the indicated cell lines after continuous treatment for 72 hours with the indicated drugs. Cellular ATP concentration was used to measure cell viability. The viability of untreated cells was set as 100%. Error bars represent the S.D. (n = 3). *CI = 0.1–0.3 and **CI < 0.1 are indicated as strong and very strong synergy between the two treatments, respectively (see Fig. 6 and Supplemental Table 1). Viability curves of wild-type (upper panels) and PARP1−/− cells (lower panels) treated with temozolomide alone or with the indicated concentrations of olaparib (A) or veliparib (B). The concentrations of PARP inhibitors are shown beside each curve in micromolar units. (C–F) Cell cycle analyses of wild-type (C and D) and PARP1−/− cells (E and F) 12 hours after the indicated drug treatments [temozolomide (TMZ; 100 μM), olaparib (1 μM), and veliparib (1 μM)]. Representative data are shown from independent experiments with consistent results experiments (C and E). Percentages of cells in the sub-G1, G1, S, and G2-M phases are shown (D and F). Total counts within the outer frame of (C) or (E) are set as 100%. Results are the average of three independent experiments. Statistical analyses were performed for the G2-M population. *P < 0.05; **P < 0.01. Data of average and S.D. for all phases are shown in Supplemental Table 2. BrdU, bromodeoxyuridine; FITC, fluorescein isothiocyanate.
To further document the marked difference between olaparib and veliparib in the cell viability assays, we examined cell cycle after drug treatments (Fig. 1, C–F). Whereas 1 μM olaparib or veliparib or 100 μM temozolomide alone had no significant impact, the combination of temozolomide with olaparib induced substantial G2-M accumulation in wild-type cells (Fig. 1, C and D; Supplemental Table 2). Temozolomide with veliparib (Fig. 1, C and D) and temozolomide in PARP1−/− cells (Fig. 1, E and F) also induced significant G2-M accumulation. However, these effects were not as pronounced as for temozolomide with olaparib. Together, these results suggest that the combination of temozolomide with olaparib (≥0.1 μM) or veliparib (≥10 μM) induces additional cytotoxicity to PARP catalytic inhibition.
Olaparib and Veliparib Potentiate Camptothecin Comparably.
Synergism between camptothecin and PARP inhibitors is well established (Smith et al., 2005; Daniel et al., 2009; Rouleau et al., 2010; Zhang et al., 2011; Brenner et al., 2012). As expected, PARP1−/− cells were hypersensitive to camptothecin (Hochegger et al., 2006) (Fig. 2, A and B, bottom panels), indicating the involvement of PARP1 in the repair of camptothecin-induced lesions. The combination of camptothecin with veliparib or olaparib synergistically sensitized wild-type DT40 cells with low CI (<0.3; Supplemental Table 1) at several points (upper panels of Fig. 2, A and B). Olaparib was more potent than veliparib. However, its concentration-dependent effect was not as pronounced as for the combination of temozolomide and PARP inhibitors (compare Fig. 2, A and B with Fig. 1, A and B). Furthermore, the sensitivity curves of both combinations never went beyond those of PARP1−/− cells (Fig. 2, A and B). Cell cycle analyses showed that the combination of 20 nM camptothecin with 1 μM olaparib or veliparib induced substantial G2-M accumulation (Fig. 2, C and D; Supplemental Table 2). Notably, this pattern was similar to that of PARP1−/− cells treated with 20 nM camptothecin (Fig. 2, E and F), suggesting that catalytic PARP inhibition causes the synergistic effect in the case of camptothecin.

Olaparib and veliparib augment the cytotoxicity of camptothecin (CPT) comparably. (A and B) Viability curves are shown as in Fig. 1A. *CI 0.1–0.3 and **CI < 0.1 are described as strong and very strong synergy between the two treatments, respectively. (C–F) Cell cycle analyses of wild-type (C and D) and PARP1−/− cells (E and F) 12 hours after the indicated drug treatments [CPT (20 nM), olaparib (1 μM), and veliparib (1 μM)]. Representative data are shown from multiple experiments (C and E). Percentages of cells in the sub-G1, G1, S, and G2-M phases are shown (D and F). Total counts within the outer frame of (C) or (E) are set as 100%. Results are the average of three independent experiments. Statistical analyses were performed for the G2-M population. *P < 0.05; **P < 0.01. Data of average and S.D. for all phases are shown in Supplemental Table 2. BrdU, bromodeoxyuridine; FITC, fluorescein isothiocyanate.
We also examined the different synergistic patterns in human prostate DU145 and glioblastoma SF295 cell lines. We compared the effect of olaparib and veliparib at 1 μM, where PAR levels are indistinguishably low (Supplemental Fig. 1, right) and viability is not affected (see the starting points at the y-axis of each curve; Fig. 3). The synergistic effect was markedly greater for olaparib with temozolomide than for veliparib with temozolomide in both cells (Fig. 3A). For camptothecin, olaparib was still more potent than veliparib in DU145, but the difference was not as pronounced as for temozolomide (Fig. 3B, top panel). Furthermore, olaparib and veliparib were comparably potent for camptothecin in SF295 cells (Fig. 3B, bottom panel). These results are consistent to those obtained with DT40 cells (Figs. 1 and 2).

Differential effect of olaparib and veliparib with temozolomide (A) or camptothecin (B) in human cell lines. Viability curves of human prostate cancer DU145 cells (top panels) and human glioblastoma SF295 cells (bottom panels) after continuous treatment for 72 hours with the indicated drug treatments [olaparib (1 μM) and veliparib (1 μM)]. Viability curves are shown as in Fig. 1A. *CI = 0.1–0.3 and **CI < 0.1 are described as strong and very strong synergy between the two treatments, respectively.
PARP Trapping by Temozolomide but Not Camptothecin in Combination with PARP Inhibitors.
We examined PARP-DNA complexes by Western blotting under different conditions (Fig. 4). Ten micromolar veliparib with temozolomide induced PARP-DNA complexes less efficiently than 1 μM olaparib in DT40 and DU145 cells (Fig. 4A, compare lanes 5 versus 8, and 13 versus 16), suggesting that a 10-fold higher concentration of veliparib than olaparib is not enough to induce the same amount of PARP-DNA complexes induced by olaparib (Fig. 4B). On the other hand, PAR levels were quite low under conditions where the differential PARP-DNA complexes were detected (Fig. 4A, lanes 5–8 and 13–16). We also confirmed that PAR levels at 1 μM olaparib and 1 μM veliparib were 1.5 and 6.6%, respectively, for DT40 cells, and 7.4 and 8.1%, respectively, for DU145 cells (Supplemental Fig. 1). Thus, olaparib and veliparib had similar PARP catalytic inhibition potencies at ≥1 μM. These results suggest that PARP inhibitors synergize temozolomide primarily by trapping PARP rather than by inhibiting the catalytic activity of PARP. Therefore, we conclude that highly potent PARP-trapping agents potentiate temozolomide more efficiently than simple catalytic PARP inhibitors, and that olaparib is superior to veliparib in combination with temozolomide.

PARP inhibitors induce PARP-DNA complexes with temozolomide (TMZ) but not with camptothecin (CPT). Western blot of chromatin-bound fractions against anti-PARP1, anti–histone H3, and anti-PAR antibodies (A). Western blot of chromatin-bound fractions against anti-PARP1 and anti-PCNA antibodies (C). Samples were prepared from wild-type DT40 cells (left) and DU145 cells (right) treated for 30 minutes and 4 hours, respectively, with the indicated drugs. Controls without drug are shown in lanes 1 and 9. Histone H3 (A) and PCNA (C) were used as positive markers for loading control. The blots are representatives of multiple experiments. (B) Quantification of PARP-DNA complexes after the indicated treatments. Signal intensity was quantified using ImageJ software (National Institutes of Health) from four independent Western blot analyses (two blots of wild-type DT40 and two blots of DU145 cells). The intensity of the PARP1 blot divided by the intensity of the corresponding histone H3 blot was measured for each treatment, and normalized to the sample of 1 μM veliparib + 1 mM TMZ treatment. Means ± S.D. (n = 4) are shown.
We also checked PARP trapping by the olaparib-camptothecin combination, and found that olaparib did not produce detectable PARP-DNA complexes in combination with camptothecin (Fig. 4C, lanes 5–7 and 12–13). These results indicate that the synergistic effect of camptothecin with PARP inhibitors is due to catalytic PARP inhibition rather than PARP trapping (Das et al., 2014). Therefore, we conclude that catalytic PARP inhibition is critical to potentiate camptothecin, and that veliparib and olaparib are both potent in combination with camptothecins.
Neither PARP Inactivation nor PARP Trapping Appears Relevant for the Combination of Olaparib with Cisplatin or Etoposide.
Next, we examined whether PARP inactivation or trapping affected the cytotoxicity of cisplatin or etoposide (Fig. 5). Whereas PARP1−/− DT40 cells were hypersensitive to temozolomide and camptothecin (Figs. 1A and 2A), PARP1−/− cells were not sensitive to cisplatin or etoposide compared with wild-type cells (Fig. 5, A and C, top panels), indicating that PARP1 is not involved in the repair of cisplatin- and etoposide-induced lesions in cellular models. Addition of olaparib in wild-type DT40 cells reduced the viability in a concentration-dependent manner (Fig. 5, A and C, bottom panels). However, the curves were almost parallel, indicating that olaparib added its own cytotoxicity to that of cisplatin or etoposide. Furthermore, the CI of each point was never below 0.3 (Supplemental Table 1), indicating a lack of strong synergy observed with temozolomide and camptothecin. Olaparib did not potentiate the activity of cisplatin and etoposide in DU145 cells (Fig. 5, B and D) to the extent observed for temozolomide and camptothecin (see Fig. 3). Furthermore, the combination of cisplatin and olaparib did not induce detectable PARP-DNA complexes (Supplemental Fig. 2). Hence, the rationale for combining PARP inhibitors with cisplatin or etoposide is based on neither PARP-DNA trapping nor PARP catalytic inhibition.

PARP1 is not involved in the repair of cisplatin- and etoposide-induced lesions. Viability curves are shown as in Fig. 1A. (A and C) Viability curves of wild-type and PARP1−/− DT40 cells treated with cisplatin (left) or etoposide (right) (top panels). (Bottom panels) Viability curves of wild-type DT40 cells in combinations with the indicated concentrations of olaparib (micromolar units beside curves) with cisplatin (left) or etoposide (right). (B and D) Viability curves of human prostate cancer DU145 cells in combination with the indicated concentrations of olaparib (micromolar units beside curves) with cisplatin (left) or etoposide (right).
Finally, to compare the drug combination results better, we prepared fraction-affected (Fa)–CI plots from the wild-type DT40 data with olaparib and/or veliparib (Fig. 6) (Chou, 2010). In these plots, we fixed the concentration of PARP inhibitors at 1 μM, because this concentration was common to all the combination studies, and because 1 μM of either olaparib or veliparib reduced PAR levels below 10%. Full information for CI is shown in Supplemental Table 1. Fa = 1.0 means 100% reduction of viability, and the CI at the highest Fa is the most meaningful for combination therapy. At the highest Fa, the combination of temozolomide with olaparib showed the lowest CI (CI = 0.046), followed by camptothecin with olaparib (CI = 0.211), camptothecin with veliparib (CI = 0.296), temozolomide with veliparib (CI = 0.357), etoposide with olaparib (CI = 0.536), and cisplatin with olaparib (CI = 0.562). All together, the combination demonstrates the highest synergisms are demonstrated for combinations of olaparib with temozolomide and for veliparib or olaparib with camptothecin.

Quantitative analyses of synergistic effects in the different combinations. (A) Fa-CI plots obtained from the data of Fig. 1 (A and B) for temozolomide in combination with 1 μM olaparib or 1 μM veliparib. (B) Fa-CI plots obtained from the data of Fig. 2 (A and B) for camptothecin in combination with 1 μM olaparib or 1 μM veliparib. (C and D) Fa-CI plots obtained from the data of Fig. 5A (bottom panel) for cisplatin (C) and Fig. 5C (bottom panel) for etoposide (D) in combination with 1 μM olaparib. (A–D) Shading reflects the level of synergism. CI between 0.3 and 0.7, CI between 0.1 and 0.3, and CI less than 0.1 indicate synergy, strong synergy, and very strong synergy, respectively. All data for CI are shown in Supplemental Table 1.
Discussion
Although combining PARP inhibitors with a wide range of drugs is actively being pursued, it is critical to elucidate whether different PARP inhibitors should be considered equal and how such combinations work because some PARP inhibitors selectively induce PARP-DNA trapping in addition to catalytic PARP inhibition (Murai et al., 2012b, 2014). Our study demonstrates marked differences among combinations of four established chemotherapeutic agent classes (temozolomide, camptothecin, cisplatin, and etoposide) with two PARP inhibitors in advanced clinical development, olaparib and veliparib, which differ by their PARP-trapping efficiencies (Murai et al., 2012b; present study).
Temozolomide induces base damage (Newlands et al., 1997), generating abasic sites that are cleaved by apurinic/apyrimidinic endonuclease 1, producing a 1-nucleotide gap with 3′-OH and 5′-deoxyribose phosphate (5′-dRP) groups at the ends of the breaks (Hazra et al., 2007). Camptothecins also induce single-strand breaks, but their 3′-DNA ends are covalently attached to topoisomerase I, whereas the 5′-DNA ends bear a sugar hydroxyl (Pommier, 2012, 2013). The binding of PARP1 depends on the DNA substrate (Horton and Wilson, 2013). PARP1 preferentially binds directly to base excision repair–intermediates with a 5′-dRP rather than to 5′-phosphate ends (Cistulli et al., 2004). This can explain the formation of PARP-DNA complexes at SSBs induced by temozolomide and suggests that combining temozolomide with a potent PARP-trapping agent, such as olaparib, is more rational than combining it with a potent catalytic inhibitor with lower PARP-trapping potency, such as veliparib (Table 1).
Summary of the differential effects of the PARP inhibitors, olaparib and veliparib, in combination with temozolomide or camptothecin
On the other hand, in the case of camptothecin, the absence of 5′-dRP ends and the steric hindrance of the breaks by the covalently bound topoisomerase I at the 3′ ends of the broken DNA (Pommier, 2012, 2013) probably explain the lack of detectable PARP-DNA complexes (Fig. 4). However, PARP is critical for the repair of topoisomerase I cleavage complexes (Smith et al., 2005; Zhang et al., 2011; Brenner et al., 2012). PARylation, which can be readily detected in camptothecin-treated cells (Zhang et al., 2011; Brenner et al., 2012), can reverse topoisomerase I cleavage complexes (Malanga and Althaus, 2004), limit replication fork collisions (Ray Chaudhuri et al., 2012; Berti et al., 2013), and facilitate homologous recombination at replication forks stalled by topoisomerase I cleavage complexes (Sugimura et al., 2008). PARP also repairs transcription-mediated DNA damage by topoisomerase I cleavage complexes by acting in the same pathway as tyrosyl-DNA phosphodiesterase 1, a critical repair enzyme that removes topoisomerase I–DNA covalent complexes (Zhang et al., 2011). We recently revealed the coupling of tyrosyl-DNA phosphodiesterase 1 and PARP1 for the repair of topoisomerase I–DNA covalent complexes (Das et al., 2014). Considering that PARP-DNA complexes are undetectable even at high concentrations of camptothecin (1 μM), and that the combination of olaparib or veliparib never exceeds the hypersensitivity of PARP1−/− cells, we conclude that the contribution, if any, of PARP-DNA complexes is minimal in the case of camptothecin. Ten micromolar veliparib, which reduces PAR levels to 1% (Supplemental Fig. 1), still was not enough to reach the hypersensitivity of PARP1−/− cells (Fig. 2B). The greater sensitivity of the PARP1−/− cell compared with the drug-treated wild-type cells could be due to the low-level residual PARylation, which might be sufficient to counter DNA damage. Nevertheless, since catalytic PARP inhibition explains the potentiation of camptothecins, highly potent PARP catalytic inhibitors devoid of cytotoxic effects, such as veliparib, can be viewed as a rational combination with camptothecins (Table 1). Indeed, recent publications show synergy between topoisomerase I inhibitors and veliparib in chemo-resistant colon cancer cell lines (in vitro and/or in vivo studies) (Zhang et al., 2011; Davidson et al., 2013; Shelton et al., 2013).
Enhanced effects of PARP inhibitors with cisplatin have been reported (Rottenberg et al., 2008; Hastak et al., 2010; Chuang et al., 2012). However, the synergistic effects in these studies are relatively weak compared with those in the reports dealing with temozolomide (Brenner et al., 2012; Kedar et al., 2012; Horton and Wilson, 2013) and camptothecin (Smith et al., 2005; Daniel et al., 2009; Zhang et al., 2011). The weaker synergistic effect of PARP inhibitors with cisplatin than with temozolomide or camptothecin would be caused by the lack of involvement of PARP in cisplatin sensitivity (Fig. 5A, top panel). The same holds true for the topoisomerase II–targeted drug etoposide. Accordingly, drug-induced topoisomerase II–DNA complexes fail to activate PARP (Zwelling et al., 1982). The lack of synergy between etoposide and PARP inhibitors has also been previously reported (Bowman et al., 2001).
In summary, we propose that combination strategies with PARP inhibitors and the choice of PARP inhibitor should be based not only on whether PARP is involved in the repair of the DNA lesions produced by the agent with which the PARP inhibitor will be combined but also on whether the lesions produced by the combination trap PARP-DNA complexes. Accordingly, PARP inhibitors that most efficiently trap PARP-DNA complexes, such as olaparib, niraparib, and BMN 673 (Murai et al., 2012b, 2014), would be preferable to veliparib in the case of temozolomide combinations. On the other hand, veliparib should remain a valuable agent in combination with camptothecins, as synergism for topoisomerase I inhibitors involves catalytic inhibition but not PARP trapping (Das et al., 2014). Table 1 summarizes these differences.
Acknowledgments
The authors thank Dr. Yasushi Okuno (Kyoto University, Japan) and Dr. Masahiko Nakatsui (Kobe University, Japan) for advising on combination effect analyses.
Authorship Contributions
Participated in research design: Murai, Ji, Pommier.
Conducted experiments: Murai, Zhang, Ji.
Contributed new reagents or analytic tools: Zhang, Ji, Morris, Takeda, Doroshow.
Performed data analysis: Murai, Zhang, Ji, Pommier.
Wrote or contributed to the writing of the manuscript: Murai, Pommier.
Footnotes
- Received September 30, 2013.
- Accepted March 10, 2014.
J.M. is a recipient of fellowships from the John Mung program (Kyoto University) and the Kyoto University Foundation. This work was supported by the Intramural Research Program of the National Institutes of Health [National Cancer Institute]; and the Center for Cancer Research [Grant Z01 BC 006150]. J.M. was supported by Japan Society for the Promotion of Science KAKENHI program [Grant 25740016]. J.J. was supported by the National Institutes of Health National Cancer Institute [Contract no. HHSN261200800001E].
↵
 This article has supplemental material available at jpet.aspetjournals.org.
This article has supplemental material available at jpet.aspetjournals.org.

Abbreviations
- ABT-888
- veliparib
- AZD-2281
- olaparib
- CI
- combination index
- dRP
- deoxyribose phosphate
- Fa
- fraction affected
- MK-4827
- niraparib
- MMS
- methyl methanesulfonate
- PAR
- poly(ADP-ribose)
- PARP
- poly(ADP-ribose) polymerase
- PCNA
- proliferating cell nuclear antigen
- SSB
- single-strand break
- U.S. Government work not protected by U.S. copyright





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}