Article Text

Abstract
Background and Aims Increased efficiency of energy harvest, due to alterations in the gut microbiota (increased Firmicutes and decreased Bacteroidetes), has been implicated in obesity in mice and humans. However, a causal relationship is unproven and contributory variables include diet, genetics and age. Therefore, we explored the effect of a high-fat (HF) diet and genetically determined obesity (ob/ob) for changes in microbiota and energy harvesting capacity over time.
Methods Seven-week-old male ob/ob mice were fed a low-fat diet and wild-type mice were fed either a low-fat diet or a HF-diet for 8 weeks (n=8/group). They were assessed at 7, 11 and 15 weeks of age for: fat and lean body mass (by NMR); faecal and caecal short-chain fatty acids (SCFA, by gas chromatography); faecal energy content (by bomb calorimetry) and microbial composition (by metagenomic pyrosequencing).
Results A progressive increase in Firmicutes was confirmed in both HF-fed and ob/ob mice reaching statistical significance in the former, but this phylum was unchanged over time in the lean controls. Reductions in Bacteroidetes were also found in ob/ob mice. However, changes in the microbiota were dissociated from markers of energy harvest. Thus, although the faecal energy in the ob/ob mice was significantly decreased at 7 weeks, and caecal SCFA increased, these did not persist and faecal acetate diminished over time in both ob/ob and HF-fed mice, but not in lean controls. Furthermore, the proportion of the major phyla did not correlate with energy harvest markers.
Conclusion The relationship between the microbial composition and energy harvesting capacity is more complex than previously considered. While compositional changes in the faecal microbiota were confirmed, this was primarily a feature of high-fat feeding rather than genetically induced obesity. In addition, changes in the proportions of the major phyla were unrelated to markers of energy harvest which changed over time. The possibility of microbial adaptation to diet and time should be considered in future studies.
- Microbiota
- diet
- obesity
- mouse models
- bacterial interactions
- diet
- nutrient absorption
- obesity
- small intestine
Statistics from Altmetric.com
- Microbiota
- diet
- obesity
- mouse models
- bacterial interactions
- diet
- nutrient absorption
- obesity
- small intestine
Significance of this study
What is already known about this subject?
-
Increased efficiency of energy harvest, due to alterations in the gut microbiota (increased Firmicutes and decreased Bacteroidetes) has been implicated in obesity in mice and humans.
-
Studies in humans have provided variable findings with respect to changes in the gut microbiota in obese individuals and placed less emphasis on changes in the proportions of Firmicutes and Bacteroidetes in obesity.
-
Recent studies in mice suggest that diet plays an important role in modulating the gut microbiota.
-
Further mouse studies have highlighted the role of the gut microbiota in the regulation of energy homeostasis, in the pathogenesis of insulin resistance, in fatty liver, in lipid and amino acid metabolism and as a modulator of host fatty acid composition.
What are the new findings?
-
While compositional changes in the faecal microbiota were confirmed, this was primarily a feature of high-fat feeding rather than genetically induced obesity.
-
The relationship between the microbial composition and energy harvesting capacity is more complex than previously considered.
-
The changes in proportions of Firmicutes and Bacteroidetes were unrelated to markers of energy harvest which changed over time.
-
The mouse gut microbiota was dominated not only by Firmicutes and Bacteroidetes, but also by a third major phylum, the Actinobacteria.
How might it impact on clinical practice in the foreseeable future?
-
Our findings suggest that microbial adaptation to diet over time, and perhaps with age, is an important variable in the complex relationship between the composition of the microbiota, energy harvesting capacity and obesity and should be taken into account in the design and interpretation of future clinical studies.
Introduction
Obesity, one of the great pandemics of our time, is a major threat to public health and to healthcare resources. This complex syndrome is influenced by host susceptibility and by environmental or lifestyle factors, such as diet and sedentary behaviour. At one level, obesity is frequently thought as the outcome of a relative imbalance in energy intake versus energy expenditure. Recently, enhanced energy harvest from dietary intake, due to an alteration in microbial composition, has been highlighted as a potential contributor to the pathogenesis of obesity.1–5
Sequencing of the gut microbiota of the caecum of genetically obese ob/ob mice revealed a 50% lower relative abundance of Bacteroidetes, whereas the Firmicutes were correspondingly higher.1 Similarly, the provision of a high-calorie, high-fat/simple carbohydrate, obesity-inducing ‘Western’ diet to wild-type mice brought about an overall decrease in the diversity of the gut microbiota, a decrease in Bacteroidetes and a bloom of a single class of Firmicutes – the Mollicutes. 3 On the basis of these investigations it was suggested that the obese microbiome possesses metabolic pathways that are highly efficient at extracting energy from food.3 ,4 This has been supported by the observation that transplantation of the microbiota of chow-fed ob/ob or Western diet-fed wild-type mice into germ-free wild-type mouse recipients resulted in mice receiving an ‘obese’ microbiota gaining more fat than recipients of a ‘lean’ microbiota.3 ,4 Furthermore, Ley et al 2 showed a decrease in the Firmicutes:Bacteroidetes ratio in obese human subjects in response to weight loss following dietary intervention. In a larger follow-up study in obese and lean twins, Turnbaugh and colleagues reported that while the faecal microbiome of obese subjects had significantly lower proportions of Bacteroidetes and higher levels of Actinobacteria compared to lean individuals, there was no change in the proportion of Firmicutes. 6
Further mouse studies have highlighted the role of the gut microbiota in the regulation of energy homeostasis,7 ,8 in the pathogenesis of insulin resistance,9–13 in fatty liver,14 in lipid and amino acid metabolism15 and as a modulator of host fatty acid composition.16 These studies suggest that interventions which target the metabolic activity of the gut microbiota may have efficacy in obesity and associated disorders. However, a variety of intrinsic and environmental factors, such as age, diet and host physiology and genotype are considered to significantly affect the structure and functionality capabilities of gut microbial communities.17–20 Indeed, it has been suggested that the Firmicutes/Bacteroidetes ratio of the human microbiota changes with age.21
A recent study in mice has shown that a high-fat diet determines the composition of the gut microbiota independent of obesity.22 Furthermore, switching from a low-fat to a high-fat diet resulted in a rapid and dramatic shift in the structure of the gut microbiota in mice in a single day.23 These studies suggest that diet plays an important role in modulating the gut microbiota. In addition, a number of follow-up studies in humans have provided variable findings with respect to changes in the gut microbiota in obese individuals and placed less emphasis on changes in the proportions of Firmicutes and Bacteroidetes in obesity.5 ,6 ,24–28 Moreover, few reports have examined the composition of the gut microbiota over time. Therefore, we examined the energy harvesting capacity and composition of the gut microbiota over time in murine models in response to high-fat (HF) feeding and genetically determined obesity. The results confirm the compositional changes in the faecal microbiota, particularly in response to diet rather than genetically determined obesity, but these are unrelated to markers of energy harvest, which change over time.
Material and methods
Animals and diets
ob/ob (leptin deficient) and wild-type C57BL/6J mice were obtained from Charles River (L'arbresle, France) and housed under barrier-maintained conditions within the biological services unit, University College Cork (UCC). Mice were received at 5 weeks of age and allowed to acclimatise for 2 weeks on a low-fat diet (10% calories from fat; Research Diets, New Brunswick, New Jersey, USA; #D12450B) to ‘normalise’ the gut microbiota. Seven-week-old male ob/ob mice, wild-type controls fed a low-fat diet and wild-type mice fed a HF-diet (45% calories from fat; Research Diets; #D12451) were housed individually for 8 weeks (n=8 per group). Body weight was assessed weekly. Precautions were taken to ensure that food intake and faecal collections were performed as accurately as possible. Food intake was measured weekly taking account of residual spillage. Faecal output per mouse was measured over 48 h at age 7, 11 and 15 weeks and weights recorded. Fat and lean body mass were measured at age 15 weeks using a Minispec mq benchtop NMR spectrometer (Bruker Instruments, Rheinstetten, Germany).
Experimental design
To examine the relationship between the composition of the gut microbiota and the efficiency of energy harvest, the effects of diet and obesity on the composition and the energy harvesting capacity of the gut microbiota were determined over time (age 7, 11 and 15 weeks). The levels of short-chain fatty acids (SCFA), the major fermentation end-products and source of energy for the host, and the energy content of the faeces were used as markers of energy harvesting.
Analysis of the composition of the gut microbiota in response to diet and obesity over time
For these studies, fresh faecal pellets (n=8 per group) were collected at 7, 11 and 15 weeks of age from lean, HF-fed and ob/ob mice. As a consequence of the number of samples that required processing, the samples were stored at −80°C prior to DNA extraction. As it has been suggested that freezing may adversely affect the levels of Bacteroidetes in stored human faecal samples,25 an initial study was undertaken to determine the impact of freezing on the microbial populations of mouse faeces. Faecal pellets were collected from C57BL/J6 mice on the same day (n=20) and divided into two groups. Faecal samples in the first group were stored on ice and DNA was extracted from these fresh samples on the day of collection (n=10) while in the second group, faecal samples were stored at −80°C and DNA was extracted (n=10) after 1 month of freezing.
DNA extractions and pyrosequencing
Total metagenomic DNA was extracted from individual faecal samples using the QIAamp DNA Stool Mini Kit (Qiagen, Crawley, West Sussex, UK) coupled with an initial bead-beating step. The microbial composition of these samples was determined by pyrosequencing of 16S rRNA tags (V4 region; 239 nt long) amplified using universal 16S primers predicted to bind to 94.6% of all 16S genes; that is, the forward primer F1 (5′-AYTGGGYDTAAAGNG) and a combination of four reverse primers R1 (5′-TACCRGGGTHTCTAATCC), R2 (5′-TACCAGAGTATCTAATTC), R3 (5′-CTACDSRGGTMTCTAATC) and R4 (5′-TACNVGGGTATCTAATC) (RDP's Pyrosequencing Pipeline: http://pyro.cme.msu.edu/pyro/help.jsp). The primers incorporated the proprietary 19-mer sequences at the 5′-end to allow emulsion-based clonal amplification for the 454 pyrosequencing system. Unique molecular identifier (MID) tags were incorporated between the adaptamer and the target-specific primer sequence, to allow identification of individual sequences from pooled amplicons. Amplicons were cleaned using the AMPure purification system (Beckman Coulter, Takeley, United Kingdom) and sequenced on a 454 Genome Sequencer FLX platform (Roche Diagnostics Ltd, Burgess Hill, West Sussex, UK) according to 454 protocols. De-noising was performed using traditional techniques implemented in the RDP pyrosequencing pipeline. Reads with low quality scores (quality scores below 40) and short length (less than 150 bp for the 16S rRNA V4 region) were removed as well as reads that did not have exact matches with the primer sequence. Clustering and statistical analysis of sequence data were performed using the MOTHUR software package.29 Trimmed fasta sequences were then BLASTed30 against a previously published 16S-specific database31 using default parameters. The resulting BLAST output was parsed using MEGAN.32 MEGAN assigns reads to NCBI taxonomies by employing the Lowest Common Ancestor algorithm which assigns each RNA-tag to the lowest common ancestor in the taxonomy from a subset of the best scoring matches in the BLAST result. Bit scores were used from within MEGAN for filtering the results prior to tree construction and summarisation (absolute cut-off: BLAST bitscore 86, relative cut-off: 10% of the top hit).31
Bomb calorimetry
Faecal samples were collected over 48 h from individual mice at 7, 11 and 15 weeks of age and used for bomb calorimetry and SCFA analysis. For bomb calorimetry analysis, the samples were weighed and oven-dried at 60°C for 48 h. The energy content of the faeces was assessed with a Parr 6100 calorimeter using an 1109 semi-micro bomb (Parr Instruments & Co., Moline, Illinois, USA). The calorimeter energy equivalent factor was determined using benzoic acid standards and each sample (100 mg) was analysed in triplicate.
SCFA analysis
SCFA analysis was performed according to previously published methods.33–35 Mice were killed at 7, 11 and 15 weeks of age and the contents of the caecum from individual mice were collected for SCFA analysis. To have sufficient quantity for gas chromatography, the caecum contents were pooled with an n=3–4 per pool. SCFA were extracted from the caecal and faecal contents using 2.0 ml Milli-Q water per 0.1 g fresh weight. The solution was centrifuged for 5 min at 10 000 g to pellet bacteria and other solids. The supernatant was collected and added to 3.0 mM 2-ethylbutyric acid in formic acid, used as the internal standard. Calibration was done using standard solutions containing 10.0 mM, 8.0 mM, 6.0 mM, 4.0 mM, 2.0 mM, 1.0 mM and 0.5 mM of acetic acid, propionic acid and butyric acid. The concentration of SCFA was determined by gas chromatography using a Varian 3500 GC system, fitted with a TRB-FFAP column (30 m x 0.32 mm x 0.50 μm; Teknokroma, Barcelona, Spain) and a flame ionisation detector. Helium was supplied as the carrier gas at an initial flow rate of 1.3 ml/min. The initial oven temperature was 100°C, maintained for 0.5 min, raised to 180°C at 8°C/min and held for 1.0 min, then increased to 200°C at 20°C/min, and finally held at 200°C for 5.0 min. The temperatures of the detector and the injection port were set at 250°C and 240°C, respectively. The injected sample volume was 0.5 μL. Peaks were integrated using Varian Star Chromatography Workstation version 6.0 software. Additional phials containing standards were included in each run to maintain calibration and a cleaning injection of 1.2% formic acid was used before each analysis.
Statistical analysis
Data for all variables were normally distributed and allowed for parametric tests of significance. Data are presented as mean values with their standard errors (SEM). Statistical analysis was performed by ANOVA and the Student t test (Graph-Pad Software, San Diego, CA, USA). Correlations analysis was performed using Pearson's (r) correlation test. p<0.05 was considered as statistically significant.
Results
ob/ob and HF-fed mice gain more weight than lean mice and vary in body composition
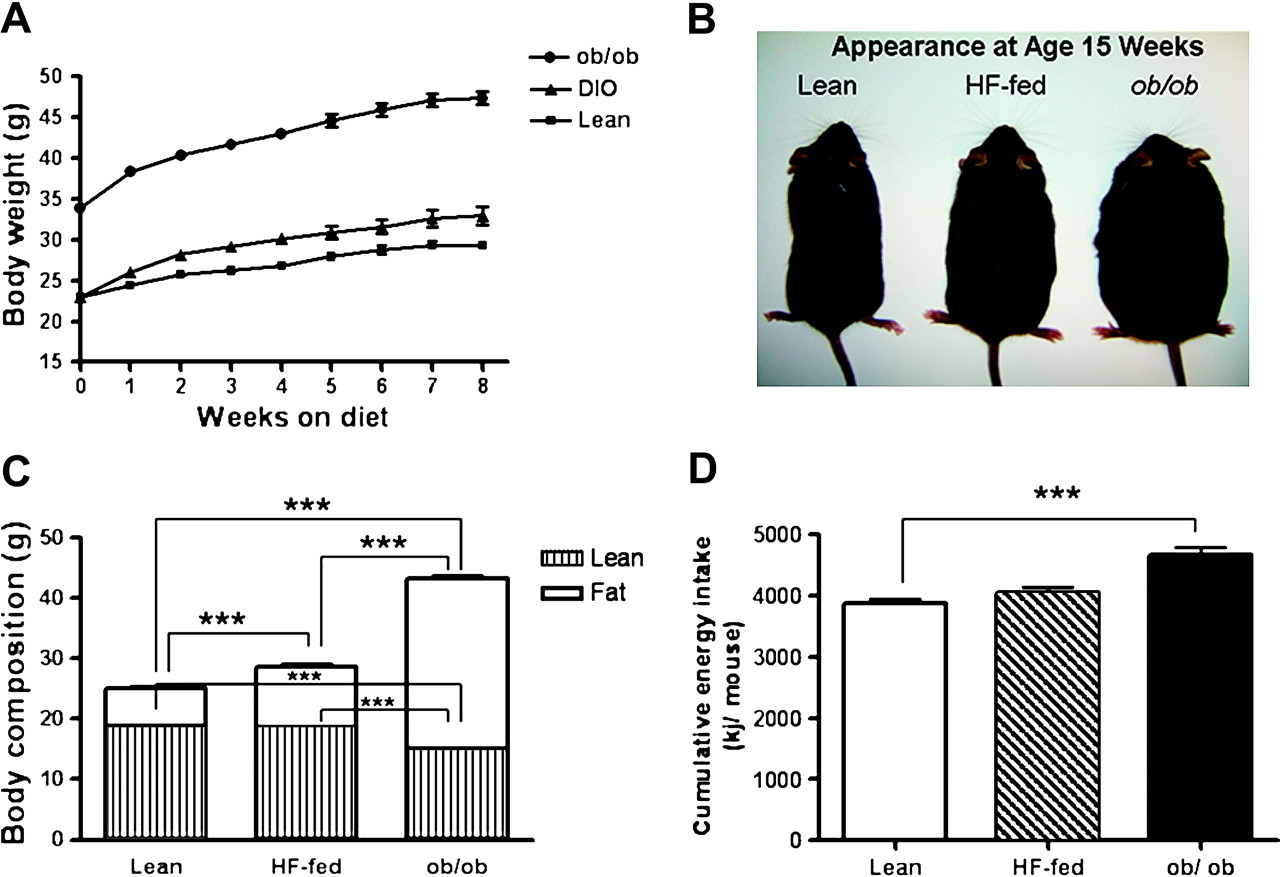
ob/ob mice gained significantly more body weight compared to lean controls (p<0.001) while a modest although statistically significant increase in body weight was observed for mice fed a high-fat diet compared to lean mice (p<0.05) over 8 weeks (figure 1A,B). The increase in body weight was associated with an increase in fat mass and a decrease in lean mass in ob/ob mice compared to lean mice, while the increase in body weight in HF-fed mice was attributable to an increase in fat mass alone (figure 1C). In agreement with previous studies, ob/ob mice consumed significantly more calories than lean controls, as measured by the cumulative caloric intake over the 8 week period of the study (figure 1D). There were no differences in cumulative caloric intake between lean and HF-fed mice.

Phenotype of mouse models of obesity. Body weight (A) and appearance (B) over the 8 week study period. Body composition (g) for lean controls, high-fat (HF) fed and ob/ob mice at age 15 weeks (C) and cumulative energy intake per mouse (kj/mouse) (D). Data presented as mean ± SEM, (n= 8 per group). ***p<0.001.
Freezing of murine faecal samples for 1 month does not significantly alter compositional analysis
It has been suggested that freezing may reduce the levels of Bacteroidetes in stored human faecal samples.25 High throughput sequencing and phylogenetic assignment of the resultant amplicons revealed that, at both the phylum and genus levels, there was no significant alteration in the composition of the gut microbiota between fresh and frozen samples (Supplementary figure 1). Therefore, for processing large numbers of samples, frozen storage was deemed acceptable prior to DNA isolation for the purposes of the present study.
supplementary files
The mouse gut microbiota is dominated by Firmicutes, Bacteroidetes and Actinobacteria
A total of 249 409 V4 16S sequence reads were generated, corresponding to an average of 31 176 reads per group or 3897 per mouse. Species richness, coverage and diversity estimations were calculated for each data set (Supplementary table 1 and Supplementary figure 2). Rarefaction curves for each group indicated that the total bacterial diversity present was well represented. Of the reads, 208 202 (83%) were assigned at the phylum level, 148 363 (59.5%) at the family level and 130 950 (52.5%) at the genus level. Taxonomy-based analysis of the assigned sequences showed that, at the phylum level, the mouse gut microbiota was dominated by Firmicutes, Bacteroidetes and Actinobacteria (figure 2 and table 1). The high proportion of Actinobacteria detected may reflect differences in the DNA extraction protocol employed or advances in primer design (primers employed match perfectly to 94.6% of sequences within RDP release 9.53 [http://pyro.cme.msu.edu/pyro/help.jsp]). Proteobacteria, Tenericutes, Spirochaetes and Deferribacteria were detected but at considerably lower levels. Consistent with the high levels of Firmicutes and Actinobacteria detected, the most dominant bacteria at the genus level were Clostridium and Bifidobacterium (table 1).

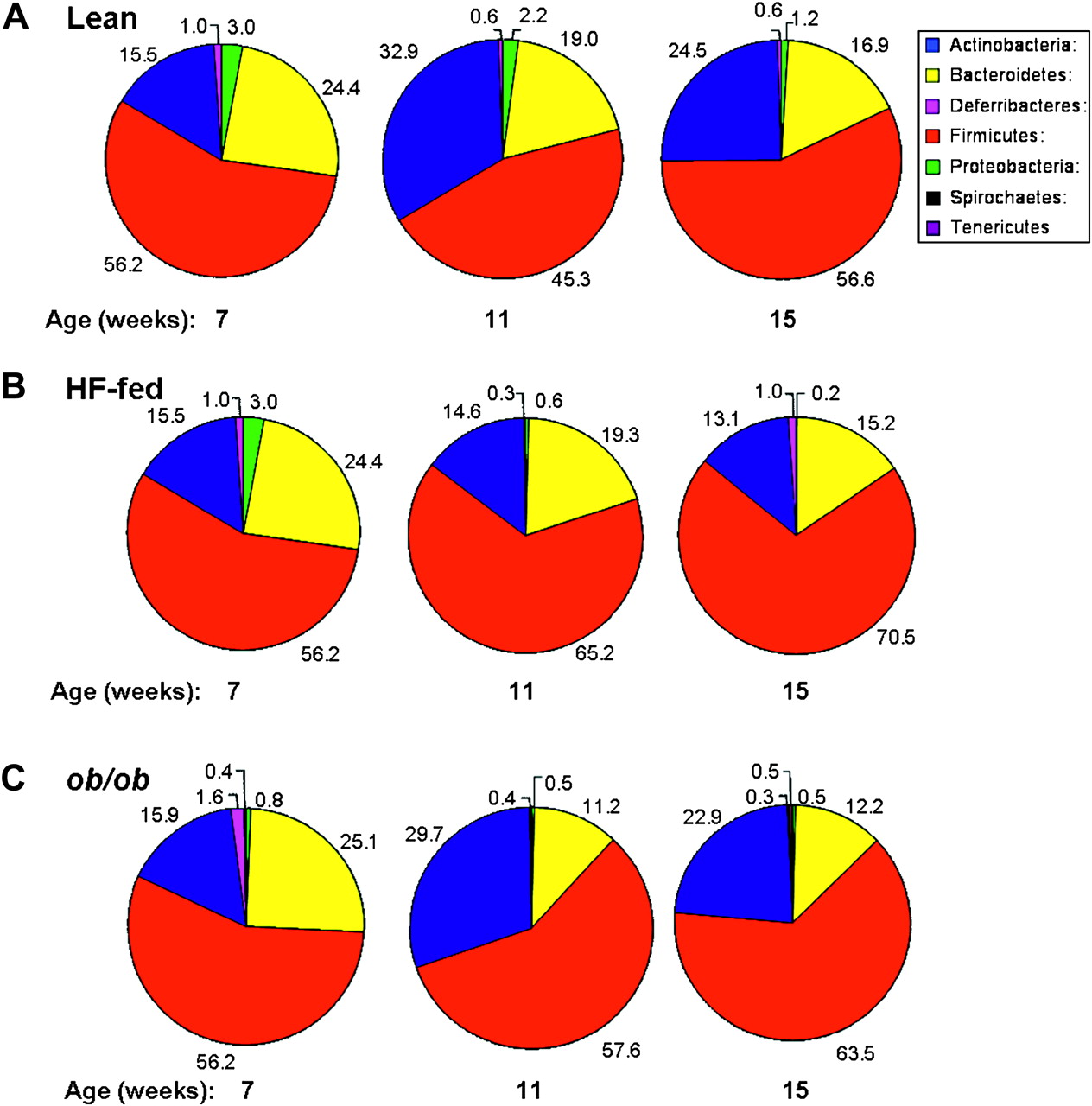
The gut microbiota composition is altered in ob/ob mice (C) at age 7, 11 and 15 weeks and in response to high-fat (HF) feeding (4 and 8 weeks) (B) but not lean mice (A) as determined by pyrosequencing of 16S rRNA tags (V4 region). Data outside the pie charts represent the mean percentage read number for the corresponding colour coded phylum (n=8 per group). Note: Microbial composition of faeces at age 7 weeks in lean mice is used as the baseline prior to introduction of HF-feeding. Thus, the pie charts for the gut microbiota composition of lean and HF-fed mice at age 7 weeks are the same only at this time-point.
Obesity and diet alter the composition of the gut microbiota in high-fat (HF)-fed and ob/ob mice over time
The composition of the murine gut microbiota changes over time in response to diet and obesity
In lean mice, from age 7 through 11 to 15 weeks, there were no significant changes in the proportions of bacteria at the level of phylum (figure 2A and table 1) or genus (table 1). In contrast, there was a progressive increase in the proportions of Firmicutes in both HF-fed and ob/ob mice, reaching statistical significance in the former (p<0.05). Reductions in Bacteroidetes over time were evident in all three groups of mice but reached statistical significance only in the ob/ob mice (p<0.001). The levels of Actinobacteria, and of the associated genus Bifidobacterium, fluctuated in all three groups with significant increases observed in ob/ob and HF-fed mice from age 7 to 11 weeks (p<0.05) but not between 7–15 or 11–15 weeks of age.
While the proportions of other phyla and genera were very low, changes were detectable. Proteobacteria were found to decrease in abundance over time in HF-fed mice (p<0.001); Deferribacteria and lactococci increased and decreased, respectively, from age 7 to 11 (p<0.001) and from 7 to 15 (p<0.001) weeks in ob/ob mice (table 1).
Energy harvesting changes in response to diet and obesity over time and does not correlate with the proportions of Firmicutes, Bacteroidetes and Actinobacteria
To examine the relationship between the composition of the gut microbiota and the efficiency of energy harvest, the levels of SCFA, the major fermentation end-products and source of energy for the host, and the energy content of the faeces were used as markers of energy harvesting. Faecal SCFA and energy content were then correlated with the levels of Firmicutes, Bacteroidetes and Actinobacteria in lean, ob/ob and HF-fed mice.
As reported by others,4 the energy content of faeces from ob/ob mice, as assessed by bomb calorimetry, was modestly but significantly lower at 7 weeks than that of lean controls (table 2 and figure 3A). However, this relationship did not persist at ages 11 and 15 weeks. In contrast, in mice on a high-fat diet, the faecal energy content increased from age 7 to 11 weeks without any further increase at 15 weeks (table 2 and figure 3A). Faecal output was similar in lean and HF-fed mice over time and there was no significant changes in faecal output between lean and HF-fed mice at age 7 (0.31 vs 0.32 g/day), 11 (0.30±0.01 vs 0.34±0.01 g/day) and 15 (0.29±0.01 vs 0.32±0.01 g/day) weeks. ob/ob mice produced significantly more faeces (0.47±0.02 vs 0.31±0.01 g/day; p<0.001) at age 7 weeks compared to lean controls. However, faecal production decreased over time in ob/ob mice from age 7 to 11 (0.47±0.02 to 0.34±0.02 g/ day; p<0.001) and from 7 to 15 (0.47±0.02 to 0.30±0.01; p<0.001) weeks of age. Calculation of energy assimilation (data not shown) suggests that the energy excreted is a function of food intake and the contribution of the gut microbiota to energy extraction is very small as suggested by Bajzer and Seeley.36
Energy harvesting capacity of the gut microbiota is altered in lean, high-fat (HF)-fed and ob/ob mice over time

{kind=link}
{kind=link}
{kind=link}
Markers of energy harvest are altered in mouse models of obesity. Faecal energy (A) and acetate (B) levels in lean and ob/ob mice at age 7, 11 and 15 weeks and in response to high-fat (HF) feeding (0, 4 and 8 weeks). Correlation of faecal energy content and faecal acetate levels in lean, HF-fed and ob/ob mice (C). Values refer to Pearson's r correlation and corresponding p value.
Faecal and caecal SCFA analysis showed that while acetate, propionate and butyrate were detected in the caecum, only acetate was detected in the faeces of the lean, HF-fed and ob/ob groups (table 2 and figure 3B). Faecal acetate concentration was higher in ob/ob compared to lean mice at age 7 and 11 weeks. However, faecal acetate levels decreased progressively over time in both the ob/ob and HF-fed mice.
Analysis of caecal SCFA showed that, at 7 weeks, caecal acetate and propionate levels were significantly higher in ob/ob mice compared to lean controls (table 2). However, this observation did not persist with time and no further pattern in caecal SCFA levels was observed in lean, HF-fed and ob/ob groups.
Correlation analysis revealed that the proportions of Firmicutes, Bacteroidetes and Actinobacteria did not correlate with energy harvesting markers (table 3). Furthermore, while a positive correlation between faecal energy content and acetate levels in lean mice was observed (r=0.50; p<0.01), there was no relationship between these parameters in ob/ob and HF-fed mice suggesting a more complex relationship between faecal energy and faecal SCFA levels in obesity (figure 3C).
The proportions of Firmicutes, Bacteroidetes and Actinobacteria do not correlate with energy harvesting markers
Discussion
The results confirmed the compositional changes in the microbiota previously linked with obesity, particularly in response to a high-fat diet, but these were unrelated to energy harvesting capacity, which changed over time. Furthermore, markers of energy harvesting were not correlated with the proportions of the major phyla. The study also showed that the mouse gut microbiota was dominated not only by Firmicutes and Bacteroidetes, but also by a third major phylum, the Actinobacteria.
This is the first report to detect high levels of the phylum Actinobacteria and the associated genus Bifidobacterium in the mouse gut microbiota using a pyrosequencing approach. There appears to be discordance in detection rates and levels of Bifidobacterium between pyrosequencing1 ,5 ,22 ,37–40 and other techniques such as fluorescence in-situ hybridisation (FISH) and traditional culture-based methods.41–43 The low percentage of Actinobacteria in some studies has been attributed to difficulties associated with extracting the associated DNA, a difference in the GC content of the template or the accuracy of PCR primers.44 In this study, the particular DNA extraction protocol employed and/or the use of primers predicted to bind to 94.6% of all 16S genes, may account for the greater proportion of Actinobacteria reads.
Another potential pitfall in studies of this type is the impact of freeze storage prior to DNA extraction.25 In our hands, a preliminary assessment revealed that storage of frozen samples over 1 month had no major influence on the DNA-based compositional analysis. It is noteworthy that a recent large metagenomics study in Europe (MetaHIT) also used frozen stool samples.45
The evidence supporting the concept that the gut microbiota in obesity facilitates the extraction of additional calories from ingested food has been reviewed elsewhere.46–48 Although we were able to confirm that the energy content of faeces from ob/ob mice was reduced and that caecal SCFA were increased at 7 weeks (consistent with increased harvest), as reported by others,4 this did not persist with time and was not found in HF-fed mice. It is noteworthy that faecal acetate levels decreased in both ob/ob and HF-fed mice over time but not in lean controls. The decrease in faecal acetate may be due to the observed alterations in the gut microbiota or an increase in its uptake/absorption in response to diet and obesity. Interestingly, it has been reported that, while obese individuals have higher faecal SCFA levels, these changes were not associated with a higher proportion of Firmicutes leading the authors to speculate that the amount of SCFA produced and not the ratio of Firmicutes and Bacteroidetes is important in obesity.24 It is unlikely that the Firmicutes:Bacteroidetes ratio is solely responsible for obesogenic shifts in the microbiota as indicated by Turnbaugh and colleagues 6 suggesting that altered proportions of other phyla including the Actinobacteria should also be further investigated. Indeed, the critical biomarker of obesity is uncertain (Firmicutes, Bacteroidetes, Firmicutes:Bacteroidetes ratio thereof and other phyla) and remains to be determined.
The primacy of diet in determining the composition of the gut microbiota independent of obesity has been elegantly demonstrated by Hildebrandt et al.22 Moreover, Turnbaugh et al, 23 demonstrated that the structure of the microbiota changes in a rapid and dramatic manner after switching from a low-fat polysaccharide-rich diet to a high-fat, high-sugar Western diet. In our study, while time-dependent decreases in Bacteroidetes in ob/ob mice were observed, the levels of Firmicutes significantly increased in HF-fed but not ob/ob mice over time. This observation suggests that a high-fat diet and not the obese genotype of ob/ob mice exerts the greater influence on the composition of the gut microbiota, in agreement with the results by Hildebrandt et al.22 Furthermore, Duncan et al,25 demonstrated a significant diet-dependent reduction in Firmicutes levels in faecal samples from obese individuals on a low-carbohydrate diet. Taken together, these observations suggest that diet plays an important role in modulating the gut microbiota and suggests that dietary variations must be taken into account in human studies.
In the present study, there was no significant change in the composition and energy harvesting of the gut microbiota of lean mice over the duration of the study suggesting that most of the changes observed in the HF-fed mice were probably attributable to change in diet. However, some temporal variations in the microbiota were apparent in the ob/ob mice over this period including a decrease over time in Bacteroidetes and Deferribacteria, at the phylum level, and Lactococcus at the genus level. While there has been an increasing appreciation of the impact of age on the gut microbial composition for example the Firmicutes/Bacteroidetes ratio of the human gut microbiota changes with age, changing from 0.4 to 10.9 to 0.6 as one moves from infants to adults and onto elderly individuals, respectively,21 we did not address age-associated alterations. However, our findings suggest that a comprehensive assessment of the role of gut microbiota in obesity should take account of temporal variation and adaptation.
Although Proteobacteria decreased in response to a high-fat diet, these bacteria were only present in low proportions in the murine gut microbiota. Interestingly, increases in the associated classes (Gamma-Proteobacteria) and family (Enterobacteriaceae) have been associated with weight loss in adolescents26 and gastric bypass in obese individuals,27 respectively.
Our data demonstrate that changes in the microbiota were dissociated from markers of energy harvest, suggesting that mechanisms other than energy harvesting may contribute to microbiota-induced alterations in obesity and metabolic diseases. In this regard, other work has suggested that the gut microbiota and its products (eg, LPS and SCFA) regulate host gene expression and, thereby, affect host energy expenditure and storage acting through mechanisms involving fasting-adipose adipose factor (FIAF),7 adenosine monophoshate (AMP)-activated protein kinase (AMPK),8 G-protein-coupled receptor (GPR)4149 and CD14/Toll-like-receptor (TLR)4.9 Indeed, germ-free mice are protected from the development of diet-induced obesity.8 Moreover, a recent study in the TLR5-deficient mice suggests that malfunction of the innate immune system may promote the development of metabolic syndrome through a mechanism involving the gut microbiota.50
In conclusion, while compositional changes in the faecal microbiota were confirmed, this was primarily a feature of high-fat feeding rather than genetically determined obesity. In addition, changes in the proportions of the major phyla of the gut microbiota were unrelated to markers of energy harvest which changed over time. These findings suggest that microbial adaptation to diet over time, and perhaps with age, is an important variable in the complex relationship between the composition of the microbiota, energy harvesting capacity and obesity and should be considered in future studies.
Acknowledgments
We thank Aileen Hogan, Andy Joyce and Talia Huffe for their technical assistance.
References
Footnotes
-
Linked articles 223594.
-
Funding The authors are supported in part by Teagasc (an Agency of the Irish Government Department of Agriculture, Fisheries and Food), by Science Foundation Ireland (in the form of a research centre grant to the Alimentary Pharmabiotic Centre and a PI award to PWOT), by Grant NIH RO1 DK058855 (to RMOD) and by Alimentary Health Ltd.
-
Competing interests None.
-
Ethics approval All animal experiments were approved by the UCC Animal Ethics Committee and experimental procedures were conducted under appropriate licence from the Irish government.
-
Provenance and peer review Not commissioned; externally peer reviewed.