Article Text

Abstract
Objective Vertical sleeve gastrectomy (VSG) produces high rates of type 2 diabetes remission; however, the mechanisms responsible remain incompletely defined. VSG increases circulating bile acid concentrations and bile acid signalling through TGR5 improves glucose homeostasis. Therefore, we investigated the role of TGR5 signalling in mediating the glucoregulatory benefits of VSG.
Design VSG or sham surgery was performed in high-fat-fed male Tgr5+/+ (wild type) and Tgr5−/− (knockout) littermates. Sham-operated mice were fed ad libitum or food restricted to match their body weight to VSG-operated mice. Body weight, food intake, energy expenditure, insulin signalling and circulating bile acid profiles were measured and oral glucose tolerance testing, islet immunohistochemistry and gut microbial profiling were performed.
Results VSG decreased food intake and body weight, increased energy expenditure and circulating bile acid concentrations, improved fasting glycaemia, glucose tolerance and glucose-stimulated insulin secretion, enhanced nutrient-stimulated glucagon-like peptide 1 secretion and produced favourable shifts in gut microbial populations in both genotypes. However, the body weight-independent improvements in fasting glycaemia, glucose tolerance, hepatic insulin signalling, hepatic inflammation and islet morphology after VSG were attenuated in Tgr5−/− relative to Tgr5+/+ mice. Furthermore, VSG produced metabolically favourable alterations in circulating bile acid profiles that were blunted in Tgr5−/− relative to Tgr5+/+ mice. TGR5-dependent regulation of hepatic Cyp8b1 expression may have contributed to TGR5-mediated shifts in the circulating bile acid pool after VSG.
Conclusions These results suggest that TGR5 contributes to the glucoregulatory benefits of VSG surgery by promoting metabolically favourable shifts in the circulating bile acid pool.
- BILE ACID
- DIABETES MELLITUS
- GASTRECTOMY
- GASTROINTESTINAL HORMONES
- COLONIC MICROFLORA
Statistics from Altmetric.com
Video abstract
Significance of this study
What is already known on this subject?
Bariatric surgery, such as vertical sleeve gastrectomy (VSG), results in high rates of type 2 diabetes remission.
Circulating bile acid concentrations are increased after VSG, which may contribute to the metabolic benefits of this surgery.
TGR5 is a bile acid receptor that contributes to the glucoregulatory effects of bile acids.
What are the new findings?
TGR5 does not contribute to the effect of VSG to reduce body weight, increase energy expenditure or increase glucose-stimulated insulin secretion.
TGR5 contributes to VSG-induced improvements in glucose regulation.
In addition to increasing circulating bile acid concentrations, VSG promotes metabolically favourable shifts in the circulating bile acid pool that rely, in part, on TGR5 signalling. Therefore, TGR5 may contribute to the glucoregulatory benefits of VSG by promoting metabolically favourable shifts in the circulating bile acid pool.
How might it impact on clinical practice in the foreseeable future?
Understanding the mechanisms driving the glucoregulatory benefits of bariatric surgery will likely improve treatment strategies for type 2 diabetes management. This work improves our understanding of the contribution of TGR5 to the metabolic benefits of VSG and reveals novel actions of TGR5 that may guide pharmaceutical targeting of this receptor.
Introduction
Bariatric surgery, such as Roux-en-Y gastric bypass (RYGB), is currently the most effective long-term treatment for obesity and often results in type 2 diabetes remission.1 ,2 However, the mechanisms by which this occurs are not well defined. Vertical sleeve gastrectomy (VSG) has gained interest as a low morbidity bariatric surgery, which is effective in producing weight loss and improving glucose regulation.3 VSG involves removal of ∼70% of the stomach by transecting the greater curvature of the stomach. Similar to RYGB, VSG produces diabetes remission and improves glucose homeostasis in both rodent and human clinical studies, making this procedure increasingly popular.3 ,4
Circulating bile acid concentrations are elevated after several types of bariatric surgery, including VSG surgery, and have been suggested to play an important role in mediating the metabolic benefits of these procedures.5 ,6 Bile acids are amphipathic steroid molecules with detergent properties that aide in digestion of dietary cholesterol and lipid. Bile acids also regulate glucose homeostasis, in part, by signalling through the transmembrane G-protein coupled bile acid receptor (TGR5).7
TGR5 is expressed in a variety of tissues and contributes to the maintenance of glucose homeostasis.7 Bile acid signalling through TGR5 has been suggested to potentiate glucagon-like peptide 1 (GLP-1) secretion from enteroendocrine L cells.8 GLP-1 exerts several antidiabetic actions including potentiation of glucose-stimulated insulin secretion (GSIS) and improved insulin sensitivity.9 In addition, in vitro studies suggest that TGR5 signalling in brown adipose tissue increases energy expenditure.10 Similar to the actions of TGR5 signalling, VSG surgery improves glucose tolerance, enhances energy expenditure and increases postprandial GLP-1 secretion.4 ,11 Furthermore, downstream TGR5 signalling components, including cyclooxygenase IV and Kir6.2, are upregulated after bariatric surgery in humans.12 Therefore, we investigated the role of TGR5 signalling in mediating the glucoregulatory benefits of VSG surgery using Tgr5+/+ and Tgr5−/− mice.
Materials and methods
Animals and diets
Heterozygous Tgr5+/− breeding pairs generated on a C57BL/6J background were obtained from Taconic Laboratories and a breeding colony established. At 1 month of age, male Tgr5−/− and Tgr5+/+ littermates were placed on a 45% energy high-fat diet (HFD) for 2 months. At 3 months of age, mice underwent sham or VSG surgery. Sham-operated animals were either fed ad libitum (S-AL) or weight matched to VSG-operated mice (S-WM) within their respective genotype (n per group: S-AL wild type (WT)=9, S-WM WT=10, VSG WT=8, S-AL knockout (KO)=11, S-WM KO=10, VSG KO=8). S-WM mice were included in order to distinguish between the body weight-dependent and body weight-independent effects of VSG. Weight-matching was performed as previously described.11 ,13 Mice were maintained on HFD throughout the study. Indirect calorimetry was performed at 5.2 months after surgery. Fasting blood samples were collected after an overnight fast at 4 months after surgery. Oral glucose tolerance tests (OGTTs) were performed at 1.5 and 3.5 months after surgery after an overnight fast (1 g/kg body weight gavage with dextrose). Fasted mice were euthanized 6 months postoperatively by an overdose of pentobarbital (200 mg/kg intraperitoneally), and tissues and cecal contents were collected. Islet immunohistochemistry, liver immunoblotting, plasma hormone measurements and bile acid profiling were performed using standard methodology (see online supplementary experimental procedures).14 ,15
VSG surgery
Mice were placed on a liquid diet (Boost, Nestlé) 4 days prior to surgery and for 14 days post surgery. Anaesthesia was induced and maintained with isoflurane (1.5–5%). A laparotomy incision was made and the stomach was isolated outside the abdominal cavity. Approximately 70% of the stomach was removed, leaving a tubular remnant. Sham surgeries were performed by isolating the stomach and placing a simple continuous pattern of suture extending through the gastric wall and along both gastric walls in the same location as the VSG-operated animals.
Gut microbial analysis
DNA was isolated from cecal contents using the MoBio PowerSoil DNA Isolation Kit (Carlsbad, California, USA). The V4 region of the 16S rRNA gene was amplified using barcoded 515F and 806R primers and sequenced on an Illumina MiSeq at the Cornell Biotechnology Resource Center Genomics Facility.16 β-Diversity was calculated using the unweighted and weighted UniFrac metrics on an OTU table rarefied to 38 000 sequences per sample.17 Taxonomic groups that were differentially abundant between treatments were identified using the Galaxy version of LEfSe.18 See online supplementary methods for further details.
Statistics and data analysis
Data are presented as mean±SEM. All data were analysed by two-factor analysis of variance with Tukey's post test, unless otherwise stated. Differences were considered significant at p<0.05.
Results
VSG surgery decreases energy intake and body weight and increases energy expenditure independent of genotype
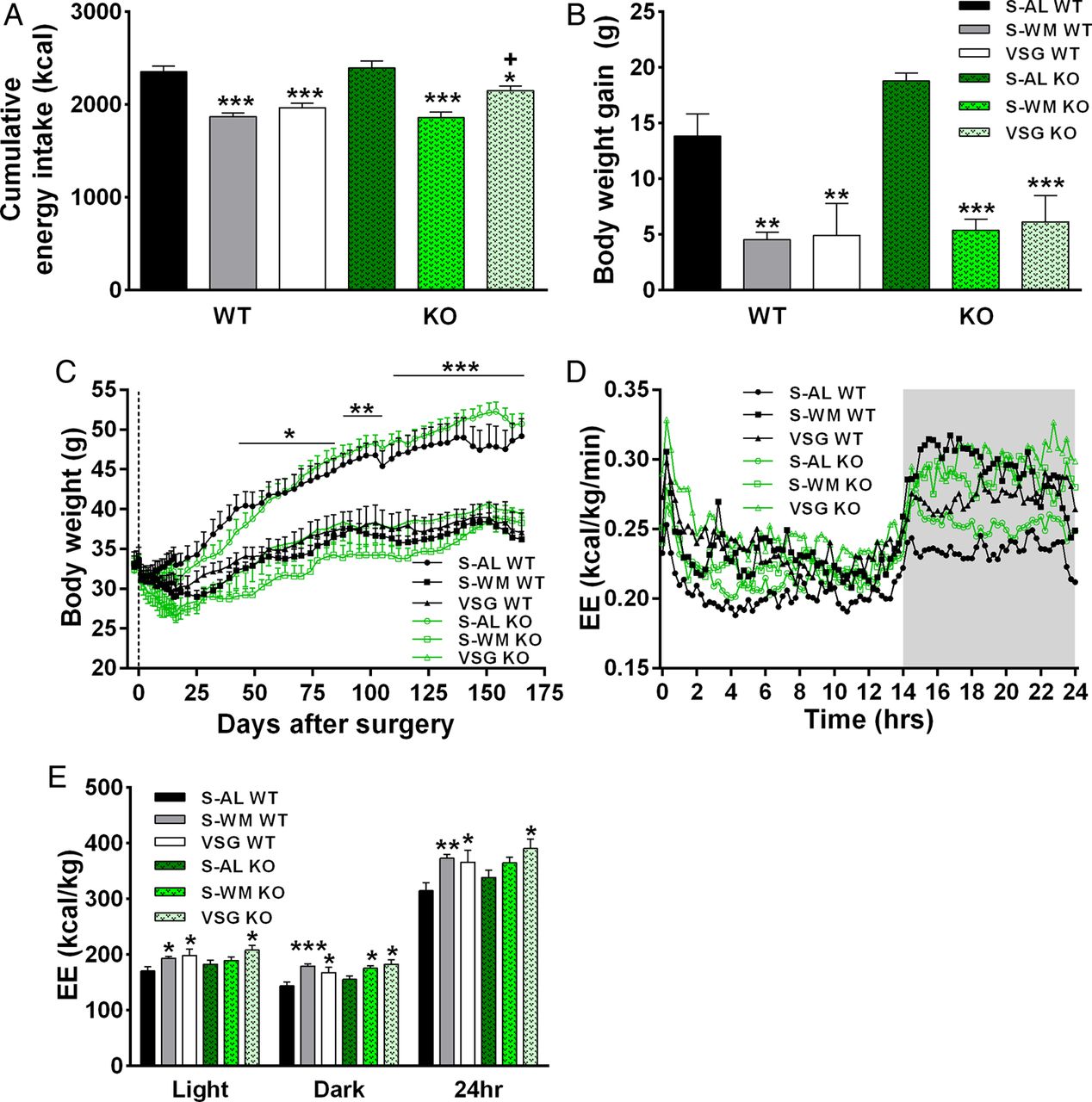
VSG surgery reduced cumulative energy intake, body weight gain and total white adipose tissue (WAT) mass in both Tgr5+/+ and Tgr5−/− mice compared with S-AL controls (figure 1A–C, see online supplementary table S1; p<0.05). Body weight and WAT mass did not differ between the S-WM and VSG-operated groups, nor did it differ between genotype for the three different treatment conditions. Therefore, we were able to assess the body weight-independent effects of VSG and body weight-independent contributions of TGR5 signalling to the glucoregulatory benefits of VSG.

TGR5 signalling does not contribute to vertical sleeve gastrectomy (VSG)-induced reductions in energy intake and body weight or postoperative increases in energy expenditure. (A) Cumulative energy intake, (B) body weight gain between days 0 and 158 post surgery, (C) body weight, (D) energy expenditure and (E) total energy expenditure. *p<0.05, **p<0.01, ***p<0.001 compared with sham-operated animals fed ad libitum (S-AL); +p<0.05 compared with sham-operated animals weight matched to VSG-operated mice (S-WM). n=8–11 per group. KO, knockout; WT, wild type.
Dietary bile acid supplementation has been reported to increase energy expenditure in vivo, with in vitro work implicating TGR5 as an upstream mediator.10 Energy expenditure did not differ between genotype within the three different treatment conditions, suggesting that TGR5 signalling does not play a crucial role in the regulation of metabolic rate in vivo. VSG resulted in an ∼15% increase in light, dark and 24 h energy expenditure compared with S-AL in both genotypes (figure 1D,E; p<0.05). Respiratory quotient did not significantly differ between groups, except for a lowering in S-WM WT compared with S-AL WT (see online supplementary table S1; p<0.05). Overall, the elevation in energy expenditure was similar between VSG WT and VSG KO mice, suggesting that TGR5 does not contribute to VSG-induced increases in energy expenditure.
The glucoregulatory benefits of VSG surgery are blunted in Tgr5−/− mice
Fasting blood glucose concentrations were reduced ∼25% in VSG WT mice compared with S-AL WT and ∼20% compared with S-WM WT at 4 months after surgery (figure 2A; p<0.05). However, fasting blood glucose in VSG KO did not significantly differ compared with S-WM KO, suggesting that VSG-induced body weight-independent improvements in fasting glycaemia are attenuated in the absence of TGR5 signalling. At 1.5 months after surgery, glucose tolerance was improved in VSG-operated mice compared with S-AL and S-WM controls in both genotypes (see online supplementary figure S1B; p<0.05). At 3.5 months after surgery, the improvement in glucose tolerance was preserved in VSG WT animals relative to S-AL WT and S-WM WT (figure 2B; p<0.05). In contrast, the glucose area under the curve (AUC) in VSG KO rose such that glucose tolerance no longer differed between VSG KO and S-WM KO. The glucose AUC decreased by –4±11% between 1.5 and 3.5 months after surgery in VSG WT, but rose by 48±25% in VSG KO (p<0.05 by Student's t test). Direct comparison of VSG WT with VSG KO revealed a 51% increase in the glucose AUC in VSG KO compared with VSG WT (see online supplementary figure S1A; p<0.05 by Student's t test). Therefore, this data set suggests that TGR5 signalling contributes to the long-term maintenance of body weight-independent improvements in glucose tolerance after VSG.

TGR5 signaling contributes to improved glucose regulation after VSG. A) Fasting blood glucose at 4 months after surgery. *p<0.05, **p<0.01 VSG vs S-AL in Tgr5+/+ and Tgr5−/−, &&&p<0.001 VSG vs S-WM in Tgr5+/+ only by two-factor ANOVA with Tukey's post-test. B) Blood glucose, C) plasma insulin and D) plasma total GLP-1 at 3.5 months after surgery during an OGTT. $p<0.05 S-WM vs S-AL in Tgr5+/+ and Tgr5−/−, **p<0.01, ***p<0.001 VSG vs S-AL in Tgr5+/+ and Tgr5−/−, +p<0.05, +++p<0.001 VSG vs S-WM in Tgr5+/+ and Tgr5−/−, &p<0.05 VSG vs S-WM in Tgr5+/+ only by two-factor ANOVA with Tukey's post-test of the AUC. n=8–11 per group. KO, knockout; WT, wild type.
The percent increase in plasma insulin concentrations from fasting to peak values was significantly higher in VSG-operated animals compared with S-AL and S-WM during both the 1.5-month and 3.5-month OGTTs, demonstrating an improvement in GSIS (see online supplementary figure S1C, figure 2C, online supplementary table S2; p<0.01). GSIS did not significantly differ between VSG WT and VSG KO at either time point, suggesting that TGR5 signalling does not contribute to postoperative increases in GSIS. Fasting plasma insulin was significantly lower in VSG WT compared with S-AL and S-WM WT during both the 1.5-month and 3.5-month OGTTs. In contrast, fasting plasma insulin did not differ among Tgr5−/− groups at 1.5 months postoperatively and was only significantly lower in VSG KO compared with S-AL KO at 3.5 months postoperatively (online supplementary table S3, p<0.05). Similarly, fasting plasma leptin and total cholesterol concentrations at 4 months after surgery were twofold to sixfold lower in VSG compared with S-AL and S-WM in Tgr5+/+ but were only reduced relative to S-AL in Tgr5−/− (see online supplementary table S3; p<0.01). Therefore, the body weight-independent reductions in fasting plasma insulin, leptin and cholesterol after VSG appear to rely, in part, on TGR5.
TGR5 signalling has been suggested to potentiate GLP-1 secretion.8 Therefore, we investigated the role that TGR5 signalling may play in the elevation of nutrient-stimulated GLP-1 secretion after VSG. GLP-1 secretion did not differ between genotype in sham-operated mice. Peak GLP-1 concentrations were approximately threefold greater in VSG-operated animals at 1.5 and 3.5 months after surgery in both genotypes (see online supplementary figure S1D, figure 2D; p<0.01). The GLP-1 AUC did not significantly differ between genotype in VSG-operated mice (see online supplementary table S2). However, the enhancement of GLP-1 secretion in VSG-operated mice relative to S-WM was mildly blunted in Tgr5−/− compared with Tgr5+/+ at 1.5 months, but not at 3.5 months, postoperatively. The GLP-1 AUC was significantly elevated in VSG WT compared with S-WM WT but only tended to be elevated in VSG KO compared with S-WM KO at 1.5 months after surgery (see online supplementary figure S1D; p<0.05). Therefore, TGR5 signalling may make a small contribution to the increase in GLP-1 secretion at early time points after VSG. Despite this difference in GLP-1 secretion, VSG-operated mice of both genotypes exhibited similar improvements in glucose tolerance during the 1.5-month OGTT. Collectively, these data suggest that the contribution of TGR5 to the long-term maintenance of improved glucose tolerance after VSG is independent of GLP-1 and insulin secretion.
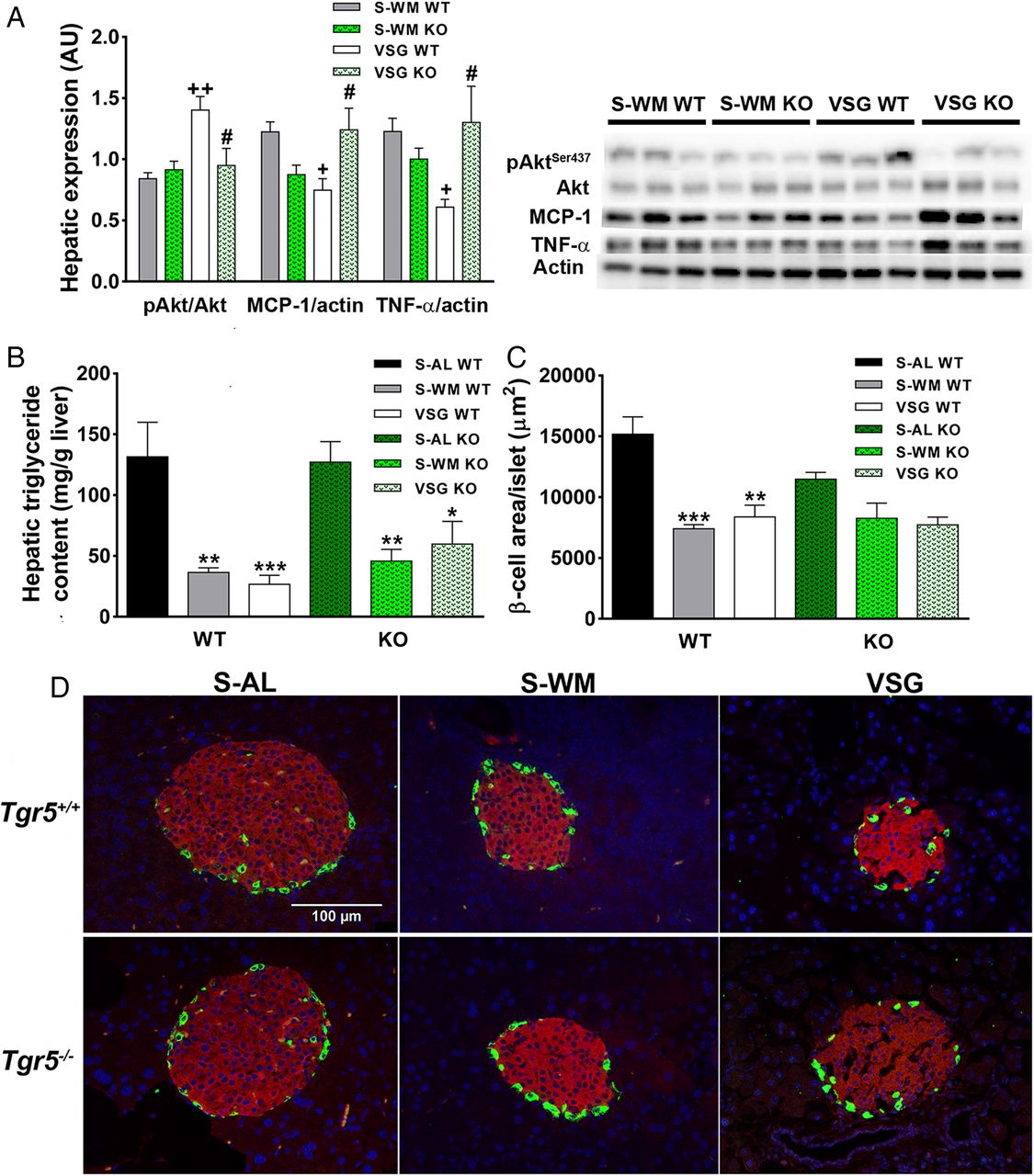
In order to investigate the molecular basis for the contribution of TGR5 to the body weight-independent enhancement in glucose tolerance after VSG, we analysed a downstream component of insulin signalling in liver of fasted animals at study termination. AktSer473 phosphorylation was ∼50% higher in VSG WT compared with S-WM WT. Furthermore, AktSer473 phosphorylation was higher in VSG WT compared with VSG KO (figure 3A; p<0.05). Two important contributors to the development of hepatic insulin resistance are increased inflammatory signalling and increased ectopic triglyceride deposition.19 Therefore, we measured hepatic triglyceride content and the protein expression of two inflammatory mediators, monocyte chemotactic protein 1 (MCP-1) and tumour necrosis factor-α (TNF-α). Hepatic triglyceride content was significantly reduced in S-WM and VSG compared with S-AL in both genotypes (figure 3B; p<0.05). Therefore, TGR5 does not appear to contribute to postoperative reductions in hepatic triglyceride deposition. VSG reduced hepatic MCP-1 and TNF-α expression relative to S-WM in Tgr5+/+, but not in Tgr5−/−. Furthermore, MCP-1 and TNF-α protein expression were elevated in VSG KO compared with VSG WT (figure 3A; p<0.05). Therefore, TGR5 may contribute to VSG-induced improvements in glucose tolerance by promoting improved hepatic insulin signalling through reductions in inflammation.

TGR5 signalling contributes to improvements in hepatic insulin and inflammatory signalling and may contribute to improvements in islet morphology after vertical sleeve gastrectomy (VSG). (A) Representative immunoblots of hepatic pAktSer437, Akt, tumour necrosis factor-alpha (TNF-α), monocyte chemotactic protein 1 (MCP-1) and β-actin. Results were quantified in densitromic units and pAktSer437 expressed relative to Akt, and TNF-α and MCP-1 expressed relative to β-actin. n=6 per group. (B) Hepatic triglyceride content. n=8–11 per group. (C) Average β-cell area per islet. *p<0.05, **p<0.01, ***p<0.001 compared with sham-operated animals fed ad libitum (S-AL); +p<0.05, ++p<0.01 compared with sham-operated animals weight matched to VSG-operated mice (S-WM); #p<0.05 compared with VSG wild type (WT). n=3–4 per group. (D) Representative images of pancreas sections immunostained for insulin and glucagon. Insulin immunostaining is in red, glucagon is in green and nuclei are stained blue. KO, knockout; WT, wild type.
Immunohistochemistry was performed on pancreatic sections collected at study termination in order to assess changes in islet morphology. Similar to our previous findings after ileal interposition surgery,14 β-cell area per islet was lower in VSG WT and S-WM WT compared with S-AL WT (figure 3C, D; p<0.001). β-Cell area per islet tended to be lower in VSG KO compared with S-AL KO, but did not significantly differ between these groups. β-Cell area per islet did not differ among the Tgr5−/− groups, suggesting that TGR5 signalling may contribute to VSG-induced improvements in islet morphology.
Metabolically beneficial shifts in the circulating bile acid profile after VSG surgery are impaired in Tgr5−/− mice
Fasting plasma bile acid profiles were assessed at 2 and 4 months after surgery in order to gain detailed insight into the effect of VSG on bile acid profile composition and the potential contribution of TGR5 to these postoperative changes. Two months after surgery, fasting plasma total bile acid concentrations were lower in S-WM WT compared with S-AL and VSG WT (see online supplementary table S4; p<0.05). At 4 months after surgery, fasting plasma total bile acid concentrations were elevated in VSG WT and VSG KO compared with their respective S-AL and S-WM control groups (figure 4A; p<0.01). Similar to our previous work in rats,11 this increase in plasma total bile acid concentrations appears to be primarily driven by an increase in conjugated bile acids (see online supplementary figure S2A, B).
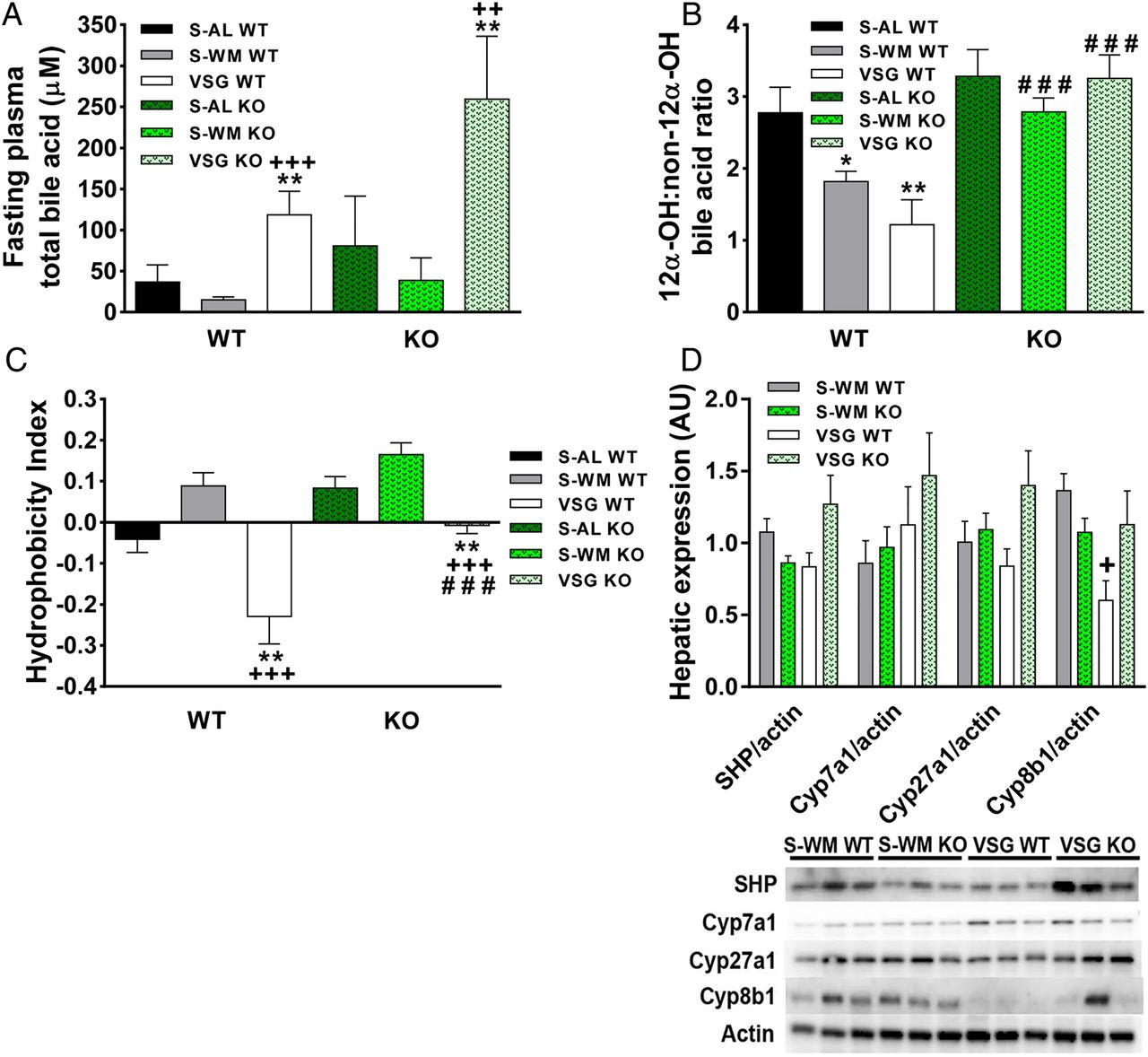
TGR5 contributes to metabolically beneficial shifts in circulating bile acid profiles after vertical sleeve gastrectomy (VSG). (A) Fasting plasma total bile acid concentration at 4 months after surgery. (B) The ratio of 12α-OH:Non-12α-OH bile acids at 4 months after surgery. (C) The hydrophobicity index of the circulating bile acid pool at 4 months after surgery. *p<0.05, **p<0.01 compared with sham-operated animals fed ad libitum (S-AL); +p<0.05, ++p<0.01, +++p<0.001 compared with sham-operated animals weight matched to VSG-operated mice (S-WM), ###p<0.001 compared with Tgr5+/+. n=8–11 per group. (D) Representative immunoblots of small heterodimer partner (SHP), Cyp7a1, Cyp27a1, Cyp8b1 and β-actin. Results were quantified in densitromic units and expressed relative to β-actin. n=6 per group. KO, knockout; WT, wild type.
In addition to increases in plasma total bile acid concentrations, it is important to take into consideration the impact of VSG-induced shifts in bile acid profiles. Increased 12α-hydroxylated bile acids (cholic acid and deoxycholic acid) to non-12α-hydroxylated bile acids (12αOH:non-12αOH) is associated with insulin resistance in humans.20 The 12αOH:non-12αOH ratio was significantly reduced in VSG WT compared with S-AL WT at 4 months after surgery, but was unchanged in VSG KO compared with S-AL and S-WM KO (figure 4B; p<0.01). Furthermore, the ratio of 12αOH:non-12αOH bile acids was twofold higher in VSG KO compared with VSG WT (figure 4B; p<0.001). Notably, regression analysis of the 3.5-month OGTT glucose AUC with the 12αOH:non-12αOH ratio revealed a positive correlation between these variables in the Tgr5+/+ groups (see online supplementary figure S2C; p<0.05, r2=0.21). At 4 months after surgery, plasma deoxycholate and taurodeoxycholate concentrations were elevated fourfold in VSG KO compared with VSG WT (see online supplementary table S4; p<0.05). Plasma taurocholate concentrations were elevated, both as an absolute value and as a percentage of the total circulating bile acid pool, in VSG KO compared with VSG WT (see online supplementary tables S4 and S5; p<0.05). These elevations in deoxycholate, taurodeoxycholate and taurocholate contributed to the increased ratio of 12αOH:non-12αOH bile acids in VSG KO.
Different bile acid subtypes exhibit varying degrees of hydrophobicity, which is determined by factors such as state of ionisation and conjugation status.21 Hydrophobic bile acids are associated with endoplasmic reticulum stress, inflammation, cell death and have been implicated in cancer pathogenesis.22 ,23 Therefore, the hydrophobicity index of the circulating bile acid pool was calculated as previously described.24 VSG resulted in a notable reduction in the hydrophobicity index in Tgr5+/+ and Tgr5−/− mice (figure 4C; p<0.001). However, the VSG-induced reduction in circulating bile acid pool hydrophobicity was blunted in Tgr5−/− compared with Tgr5+/+ (p<0.001), suggesting that TGR5 signalling contributes to this important shift in the circulating bile acid profile. TGR5-mediated increases in muricholates (hydrophilic bile acid species) and decreases in deoxycholates (hydrophobic bile acid species) contributed to the reduction in bile acid hydrophobicity index in VSG-operated Tgr5+/+ mice. Muricholates increased as a percentage of the total circulating bile acid pool in Tgr5+/+ mice, but not in Tgr5−/− mice. In particular, tauro-α-muricholate was twofold lower in VSG KO compared with VSG WT at 4 months after surgery (see online supplementary table S4; p<0.05).
In order to gain preliminary insight into how TGR5 may influence bile acid profile, we measured hepatic protein expression of small heterodimer partner (SHP), Cyp27a1, Cyp7a1 and Cyp8b1. SHP is an FXR regulated gene that acts as an atypical nuclear receptor to decrease hepatic bile acid synthesis and, therefore, could play a role in VSG-induced alterations in bile acid metabolism.21 Hepatic SHP, Cyp7a1 and Cyp27a1 protein expression did not differ between groups (figure 4D). Interestingly, hepatic Cyp8b1 protein expression was approximately twofold lower in VSG WT compared with S-WM WT and tended to be lower compared with VSG KO (figure 4D; p<0.05). Cyp8b1 is a critical enzyme in the biosynthesis of cholic acid; one of the main bile acid subtypes responsible for the increase in the 12αOH:non-12αOH ratio in VSG KO compared with VSG WT.21 Therefore, our data suggest that there is a TGR5-dependent reduction in Cyp8b1 expression after VSG that contributes to the TGR5-dependent postoperative decrease in the 12αOH:non-12αOH ratio.
Differential shifts in gut microbial populations in VSG-operated Tgr5−/− mice
Primary bile acids are synthesised in the liver and are then secreted into the gut where they are converted into secondary hydrophobic bile acids by specific gut microbes.25 Thus, alterations in gut microbial populations may contribute to shifts in bile acid profile after VSG. Therefore, we assessed gut microbial populations within cecal contents by 16S rRNA profiling. Unweighted UniFrac analysis revealed differential clustering of VSG groups compared with S-WM and S-AL groups along principal component 1 (figure 5A; p<0.001). However, weighted UniFrac analysis did not reveal differential clustering between groups (see online supplementary figure S3A), suggesting that low abundance bacteria comprise the major differences between VSG and controls. Rarefaction analysis revealed no significant differences between groups in gut microbiome richness (α-diversity) (see online supplementary figure S3B).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Vertical sleeve gastrectomy (VSG) surgery alters cecal microbial profiles. (A) Principal coordinate analysis plot of unweighted UniFrac distances, (B) relative abundance of bacterial phyla, (C) relative abundance of Akkermansia, (D) relative abundance of Turicibacteraceae, (E) relative abundance of Adlercreutzia and (F) relative abundance of Enterococcus. **p<0.01 compared with sham-operated animals fed ad libitum (S-AL), ++p<0.01 compared with sham-operated animals weight matched to VSG-operated mice (S-WM). n=8–11 per group. KO, knockout; WT, wild type.
At the phyla level, the relative abundance of members of the non-photosynthetic cyanobacteria (class Melainabacteria) was approximately 10-fold higher in VSG WT compared with S-AL WT and S-WM WT (figure 5B; p<0.01). However, the relative abundance of cyanobacteria remained low and unchanged in VSG KO compared with S-AL and S-WM KO, suggesting that TGR5 signalling contributes to this postoperative change. Verrucomicrobia is a phylum of bacteria that is typically represented in the mammalian gut by just a few genera, including the genus Akkermansia, which was decreased in VSG relative to S-AL and S-WM in both genotypes (figure 5C; p<0.01). In contrast, previous studies in rodents and humans report elevations of this genus after RYGB surgery, illustrating differential shifts in gut microbial populations in different bariatric surgery types.26 ,27 Akkermansia is known to dwell in and degrade mucus, so shifts in its abundance may reflect changes to the mucosal dynamics of the system. The relative abundance of Turicibacteraceae, a family within the Firmicutes phylum, was 20-fold lower in the VSG groups compared with the S-AL groups (figure 5D; p<0.01). Previous studies have reported decreased Firmicutes after RYGB surgery;26 ,27 however, alterations in Turicibacteraceae have not been previously reported.
At the genus level, there was a twofold decrease in the relative abundance of the genus Adlercreutzia in VSG WT and KO compared with controls (figure 5E; p<0.01). This genus is a relatively newly identified member of the phylum Actinobacteria (family Coriobacteriaceae). Actinobacteria are positively associated with obesity in lean and obese twins28 and Coriobacteriaceae are lower in human RYGB patients.27 Enterococcus was enriched in the VSG-operated groups compared with the S-AL and S-WM groups (figure 5F). Enterococcus species may be a promising probiotic with recent work suggesting that Enterococcus faecalis reduces adiposity and hepatic steatosis in high-fat-fed mice.29 Given that VSG-induced changes in Akkermansia, Adlercreutzia, Turicibacteraceae and Enterococcus were present in both genotypes, these shifts do not appear to contribute to TGR5-driven improvements in glucose homeostasis. Given that bacterial species known to play an important role in bile acid metabolism did not differ between VSG WT and VSG KO, it does not appear that alterations in the relative abundance of different gut microbial species contribute to VSG-induced shifts in bile acid profile.
Discussion
This work suggests that bile acid signalling through TGR5 is a contributor to the glucoregulatory benefits of VSG. While TGR5 does not appear to contribute to decreased energy intake or body weight gain after VSG, a lack of TGR5 signalling attenuated the body weight-independent improvements in fasting glycaemia, fasting insulinemia, hepatic insulin signalling and glucose tolerance. Bile acid signalling through TGR5 has been previously reported to contribute to glucose regulation, potentially by enhancing GLP-1 secretion and energy expenditure.7 However, TGR5 signalling does not contribute to the effect of VSG to increase energy expenditure and only mildly contributes to the effect of VSG to increase GLP-1 secretion. Instead, TGR5 signalling appears to contribute to improved glucose regulation after VSG by promoting metabolically favourable shifts in the circulating bile acid pool.
TGR5 signalling has been reported to contribute to glucose homeostasis through the potentiation of GLP-1 secretion and enhancement of energy expenditure.8 ,10 Although there was a small attenuation of postprandial GLP-1 secretion in VSG KO mice relative to VSG WT at 1.5 months after surgery, this did not appear to have a significant impact on glucose homeostasis. Notably, recent publications in rodent models report that enhanced GLP-1 receptor signalling does not play an important role in mediating the metabolic benefits of RYGB or VSG.30 ,31 Therefore, it is unlikely that TGR5-mediated potentiation of GLP-1 secretion contributes to the glucoregulatory benefits of VSG. In addition, Tgr5+/+ and Tgr5−/− mice have similar energy expenditure after sham or VSG surgery, suggesting that TGR5 signalling does not play a crucial role in the enhancement of metabolic rate.
Our data suggest that TGR5 signalling contributes to the metabolic improvements seen after VSG by promoting metabolically favourable shifts in the composition of the circulating bile acid pool. Specifically, TGR5 contributes to VSG-induced reductions in the ratio of 12αOH:non-12αOH bile acids and reductions in the circulating bile acid pool hydrophobicity index. Reductions in bile acid hydrophobicity may have contributed to VSG-induced reductions in hepatic inflammation, favouring improved insulin sensitivity23 ,32 For example, hydrophilic bile acids, such as ursodeoxycholic acid, decrease inflammation and endoplasmic reticulum stress, resulting in improved insulin sensitivity in mice.33 In contrast, hydrophobic bile acids, such as deoxycholic acid, damage cell membranes and promote inflammation and carcinogenesis in humans.22 ,34–36 Furthermore, VSG-operated mice exhibited a TGR5-dependent decrease in the ratio of 12αOH:non-12αOH bile acids. This ratio has been elegantly shown to negatively correlate with insulin sensitivity in humans.20 However, the enzyme responsible for hepatic production of cholic acid, Cyp8b1, is regulated by insulin with insulin resistance resulting in impaired inhibition of Cyp8b1, and therefore, enhanced cholic acid production.37 Whether changes in this ratio simply reflect alterations in insulin sensitivity or actively contribute to alterations in insulin sensitivity remains unknown. Interestingly, Cyp8b1 appears to be downregulated after VSG through a TGR5-dependent mechanism, suggesting that the decrease in the 12αOH:non-12αOH bile acid ratio is not simply a by-product of improved insulin sensitivity. Notably, bile acid signalling through FXR has been suggested to contribute to the effect of VSG to reduce body weight and improve glucose homeostasis,38 emphasising the important contributions made by bile acid signalling to the metabolic benefits of VSG.
Gut microbial profiling revealed a genotype-dependent increase in the relative abundance of cyanobacteria in VSG-operated mice. Therefore, TGR5 signalling may also contribute to the metabolic benefits of VSG by favouring specific shifts in gut microbial populations. Cyanobacteria have been previously found in human and rodent gut microbial populations and are associated with high-fibre diet and health.39 The pronounced genotype-dependent increase in cyanobacteria in VSG-operated Tgr5+/+ mice suggests that these gut microbes may be regulated by TGR5 signalling and could play an important metabolic role requiring further investigation. The lack of a difference in bacterial species known to play an important role in bile acid metabolism suggests that TGR5-mediated gut microbial population shifts do not contribute to VSG-induced shifts in bile acid profile. However, further metagenomic and metatranscriptomic analysis is needed in order to determine whether there are shifts in overall gut microbial bile acid enzyme gene expression.
In conclusion, this study provides novel data on the mechanisms by which bile acid signalling contributes to the metabolic benefits of VSG. Importantly, our data suggest that not only do increases in circulating total bile acid concentrations likely contribute to metabolic improvements after VSG, but that shifts in bile acid profile may be making an important contribution as well. Furthermore, this study identifies a role for TGR5 in the regulation of bile acid metabolism. TGR5-dependent decreases in hepatic Cyp8b1 expression may contribute to the effect of VSG to alter bile acid profile after VSG. However, further work is needed to fully understand how TGR5 regulates bile acid metabolism. Finally, these results emphasise the potential therapeutic utility of targeting TGR5 for the treatment of type 2 diabetes.
Acknowledgments
We thank Cornell Biotechnology Resource Center Genomics Facility for performing sequencing for gut microbial composition analysis. We thank Dr Katie Kelly for editorial input and Marco Hiller for technical support.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
- Data supplement 1 - Online supplement
Footnotes
Correction notice This article has been corrected since it published Online First. Figure 4 has been corrected.
Contributors AKM acquired and interpreted data and wrote the paper. DG, ZMH and JC acquired and interpreted data and revised the manuscript. AB, FGH and REL contributed to study design, data interpretation and revised the manuscript. MLC contributed to study design, acquired and interpreted data and revised the manuscript. BPC supervised the study, contributed to study design, data interpretation and finalised the manuscript.
Funding This research was supported by DK076169, funding from Eli Lilly, start-up support from Cornell University, the President's Council of Cornell Women and R01DK095960. BPC's laboratory also received funding during the project period from the Cornell Comparative Cancer Biology Training Program, the US Highbush Blueberry Council and U24DK092993. The FGH lab is supported by R01DK090492 and R01DK095359.
Competing interests BPC's laboratory receives funding from Eli Lilly and MLC is employed by Eli Lilly.
Ethica approval The experimental protocols were approved by the UC Davis and Cornell University Institutional Animal Care and Use Committees.
Provenance and peer review Not commissioned; externally peer reviewed.
Data sharing statement Gut microbiome data will be made publicly available through Qiime.