Article Text

Abstract
Objective Hypomethylation of LINE-1 elements has emerged as a distinguishing feature in human cancers. Limited evidence indicates that some LINE-1 elements encode an additional internal antisense promoter, and increased hypomethylation of this region may lead to inadvertent activation of evolutionarily methylation-silenced downstream genes. However, the significance of this fundamental epigenetic mechanism in colorectal cancer (CRC) has not been investigated previously.
Design We analysed tissue specimens from 77 CRC patients with matched sets of normal colonic mucosa, primary CRC tissues (PC), and liver metastasis tissues (LM). LINE-1 methylation levels were determined by quantitative bisulfite pyrosequencing. MET, RAB3IP and CHRM3 protein expression was determined by western blotting and IHC. MET proto-oncogene transcription and 5-hydroxymethylcytosine (5-hmc) were evaluated by quantitative real-time-PCR.
Results Global LINE-1 methylation levels in LM were significantly lower compared with the matched PC (PC=66.2% vs LM=63.8%; p<0.001). More importantly, we observed that specific LINE-1 sequences residing within the intronic regions of multiple proto-oncogenes, MET (p<0.001), RAB3IP (p=0.05) and CHRM3 (p=0.01), were significantly hypomethylated in LM tissues compared with corresponding matched PC. Furthermore, reduced methylation of specific LINE-1 elements within the MET gene inversely correlated with induction of MET expression in CRC metastases (R=−0.44; p<0.0001). Finally, increased 5-hmc content was associated with LINE-1 hypomethylation.
Conclusions Our results provide novel evidence that hypomethylation of specific LINE-1 elements permits inadvertent activation of methylation-silenced MET, RAB3IP and CHRM3 proto-oncogenes in CRC metastasis. Moreover, since 5-hmc content inversely correlated with LINE-1 hypomethylation in neoplastic tissues, our results provide important mechanistic insights into the fundamental processes underlying global DNA hypomethylation in human CRC.
- Colorectal Neoplasia
- Methylation
- Oncogenes
Statistics from Altmetric.com
Significance of this study
What is already known about this subject?
-
Global hypomethylation is one of the distinguishing features in human cancers.
-
Certain LINE-1 elements encode an additional internal antisense promoter which may offer an alternative transcriptional start site for specific downstream genes.
-
Limited evidence indicates that LINE-1 hypomethylation associates with activation of downstream genes in chronic myeloid leukaemia and bladder cancer; however, no previous studies have investigated the underlying mechanism of LINE-1 element hypomethylation in the development of CRC and cancer metastasis.
What are the new findings?
-
This study provides the first evidence for increased LINE-1 hypomethylation as a key feature in cancer metastasis; in particular, their significant demethylation in liver metastasis tissues of patients with colorectal cancer (CRC).
-
This study elucidates the role of hypomethylation of certain LINE-1 elements in activating several proto-oncogenes (MET, RAB3IP and CHRM3) in CRC metastasis development.
-
Overexpression of MET oncoprotein expression is primarily triggered and controlled by the hypomethylation of its internal antisense promoter located within certain LINE-1 elements, and not by its own promoter region in CRC and distant metastases tissues.
-
This study highlights that enhanced accumulation of 5-hydroxymethylcytosine (5-hmc) inversely correlates with global LINE-1 hypomethylation, providing fundamental insights into the basic processes that regulate global hypomethylation in cancer cells.
How might it impact on clinical practice in the foreseeable future?
-
Our study highlights a molecular perspective for LINE-1 hypomethylation-induced activation of multiple proto-oncogenes in CRC metastasis. Moreover, we provide unique evidence that the amount of 5-hmc correlates with LINE-1 sequence demethylation in cancer tissues. Taken together, these results underscore the potential diagnostic and prognostic value of LINE-1 hypomethylation as a biomarker in metastatic cancer.
Introduction
More than 98% of the human genome consists of transcriptionally inactive non-coding DNA elements1–3 which predominantly includes the long interspersed nuclear element-1 (LINE-1) repetitive sequences. LINE-1 sequences constitute ∼18% of the entire human genomic mass, and comprise a 5′-untranslated region (UTR), two open-reading frames, and a 3′-UTR.4–6 LINE-1 elements contain a CpG island in their 5′-UTR region which is often heavily methylated in normal somatic cells.7 ,8 Alteration in the methylation status of LINE-1 elements is considered to be a useful surrogate marker for the global DNA methylation content.8 ,9
Global loss of cytosine methylation within CpG dinucleotides is one of the distinguishing features of neoplastic cells in human cancers.10 Genomic DNA hypomethylation is associated with increased chromosomal instability and global loss of imprinting which plays a central role in tumorigenesis.11 ,12 Given the diversity and extent of genomic instability that ensues following genomic hypomethylation, some studies have suggested that global DNA hypomethylation may have an even more important role in human cancers than the hypermethylation-induced transcriptional silencing of specific tumour suppressor genes.13 ,14
Previous studies have highlighted the importance of LINE-1 hypomethylation in various human cancers, including bladder cancer,15 chronic myeloid leukaemia,16 gastric cancer,17–19 gastrointestinal stromal tumours,20 ovarian cancer21 and hepatocellular carcinoma.22 In colorectal cancer (CRC), LINE-1 hypomethylation significantly correlates with various clinical and pathological variables.23–25 Previously, we have demonstrated that LINE-1 hypomethylation is one of the distinguishing features of non-Lynch Syndrome familial CRC.23 Likewise, hypomethylation of LINE-1 inversely correlated with microsatellite instability (MSI) and the CIMP phenotype in CRC.24 Additionally, loss of LINE-1 methylation has been shown to associate with more aggressive progression of CRC.25 Nonetheless, the functional mechanisms underlying LINE-1 hypomethylation in CRC, and distant metastasis development, remain poorly understood.
A full-length LINE-1 element consists of two promoters within its 5′ UTR region that are aligned in opposing orientations.26 One of these promoters acts as a sense-promoter and regulates the transcription of retrotransposon-related genes such as an RNA-binding protein, an endonuclease, and a reverse transcriptase. The second promoter which is aligned in the opposite direction, functions as an antisense promoter, and provides an alternative transcription start site (TSS) that results in the induction transcription of downstream genes.26 In the human genome, more than 275 000 TSSs have been identified within such repetitive elements, accounting for ∼31.4% of all TSSs.27 Although a few previous studies have suggested that LINE-1-associated transcripts may be involved in chronic myeloid leukemia16 and bladder cancer,15 the precise role and regulatory mechanism of LINE-1 hypomethylation-induced oncogene activation have, thus far, not been investigated in CRC and the development of distant metastasis.
5-hydroxymethylcytosine (5-hmc) is an oxidised product of 5-methylcytosine (5-mc) which was first identified in mammals in 1972.28 The Ten-Eleven-Translocation family of oxygenases catalyses the transfer of a hydroxyl group to 5-mc resulting in the formation of 5-hmc.29 In embryonic stem cells, dynamic regulation of 5-hmc status is involved in cell renewal and differentiation.30 ,31 Impaired formation of 5-hmc is associated with myeloid tumorigenesis.32 Several recent studies have suggested that the 5-hmc is an intermediate product during catalytic conversion of 5-mc to 5-c and plays a critical role in DNA demethylation induction by inhibiting the binding of methyl-CpG binding domain (MBD) proteins and chromatin remodelling-associated complexes.33–35 However, to the best of our knowledge, no previous studies have interrogated the correlation between the presence of 5-hmc and hypomethylation of LINE-1 elements in cancer, a concept that could help in a better understanding of the role of 5-hmc and aberrant DNA methylation in cancer.
In the present study, we have addressed this important gap in knowledge, and have examined the methylation patterns of LINE-1 repetitive sequences during CRC metastasis development by directly comparing primary CRCs with matching liver metastases. Herein, we provide the first evidence for increased LINE-1 hypomethylation in cancer metastasis, particularly in LM in patients with CRC. We also demonstrate that hypomethylation of specific intronic LINE-1 elements permits inadvertent activation of multiple proto-oncogenes including MET, RAB3IP and CHRM3 in CRC and liver metastasis, suggesting a potential role of these oncogenes in enhanced malignant behaviour in colorectal neoplasia. More importantly, we noted that overexpression of MET oncoprotein was primarily triggered and controlled by the hypomethylation of its internal antisense promoter located within these unique LINE-1 elements, and not by its own promoter region in CRC metastases tissues. Finally, this study highlights that enhanced accumulation of 5-hmc inversely correlates with global LINE-1 hypomethylation, providing fundamental insights into the basic processes that regulate global hypomethylation in cancer cells.
Materials and methods
Cell lines and 5-aza-2-deoxy-cytidine treatment
Seven CRC cell lines, HCT116, RKO, SW48, Caco-2, HT29, SW480 and SW620 were obtained from the American Type Culture Collection (ATCC, Rockville, Maryland, USA). Cell lines were treated with 2.5 μM 5-Aza-2′-deoxycytidine (5-aza-dC; Sigma–Aldrich, St Louis, Missouri, USA) for 72 h, as described previously.36
Tissue specimens
A total of 77 formalin-fixed, paraffin-embedded (FFPE) matched corresponding normal colonic mucosa (NM), primary CRC tissues (PC) and liver metastasis (LM), were microdissected and analysed. CRC tissues were obtained from La Paz University Hospital, Madrid, Spain. Written informed consent was obtained from all patients, and the study was approved by the institutional review boards. Detailed demographic and clinicopathological features of these patients are shown in online supplementary table S1.
DNA methylation analysis
Methylation levels of repetitive sequences (global LINE-1 and local LINE-1) were analysed by quantitative bisulfite pyrosequencing using the PSQ HS 96A pyrosequencing system (Qiagen, Valencia, California, USA), as described previously,23 as shown in online supplementary table S2.
DNA copy number analysis
MET gene copy number variations were analysed by qBiomarker Copy Number PCR Assay (Qiagen). Specific MET gene region (chr7: 116312459—116438440) was amplified and normalised with qBiomarker Multicopy Reference Copy Number PCR Assay (Qiagen) following the manufacturer's instructions.
Gene expression analysis
Total RNA extracted from CRC cell lines and FFPE specimens was reverse transcribed to cDNA using random hexamers, as described previously.37 For quantitative real-time PCR (qRT-PCR), target gene expression was normalised to GAPDH expression, as described in online supplementary table S2.
Protein expression analysis
Western blotting was performed using anti-Met (Millipore, Billerica, Massachusetts, USA), anti-RAB3IP (Sigma–Aldrich), and anti-CHRM3 (Santa Cruz, Santa Cruz, California, USA) antibodies, as described previously.36 For immunohistochemical staining, h-Met SP44 primary antibody (Ventana, Tucson, Arizona, USA) was used together with the peroxidase-based EnVisionTM immunostaining kit (Dako Corporation, Carpinteria, California, USA). Expression intensity was scored as 0 (negative), 1 (weak positive), 2 (moderate positive) and 3 (strong positive).
5-hydroxymethylcytosine (5-hmc) detection analysis
DNA glucosylation and digestion was performed using the Quest 5-hmC Detection Kit (Zymo Research, Irvine, California, USA). A total of 400 ng of DNA was treated with 5-hmc glucosyltransferase at 37°C for 2–3 h using a 5-hmc glucosyltransferase buffer containing uridine diphosphoglucose. Glucosylated DNA was digested with MspI at 37°C for 4 h. Global LINE-1 and local LINE-1 specific 5-hmc detection and quantification were performed by qRT-PCR, as described in online supplementary table S2.
Statistical analysis
Data were analysed with GraphPad Prism V.5.0 software. To evaluate significant differences between two matched pair groups of samples, paired t test was used. Pearson's correlation coefficient (r) was used to measure correlation, and logarithmic regression was used to calculate the R2 and the slope.
Results
LINE-1 methylation is involved in CRC metastasis
The methylation status of LINE-1 repetitive sequences in HCT116, RKO, SW48, Caco-2, SW480, and SW620 human CRC cell lines was analysed by quantitative pyrosequencing (figure 1A). All microsatellite stable (MSS) cell lines were significantly hypomethylated (Caco-2=68.1%, SW480=51.8%, and SW620=39.2%; p<0.05) at LINE-1 repetitive elements compared with the CRC cell lines with MSI (HCT116=76.6%, RKO=75.5%, and SW48=74.8%). This observation was corroborated by our previous report in which we demonstrated that LINE-1 sequences were significantly hypomethylated in non-Lynch Syndrome familial CRC.23 Additionally, marked differences were observed between two CRC cell lines derived from the same patient, in which LINE-1 sequences were significantly more hypomethylated in SW620 (derived from a lymph node metastasis) compared with SW480 cells (primary tumour).
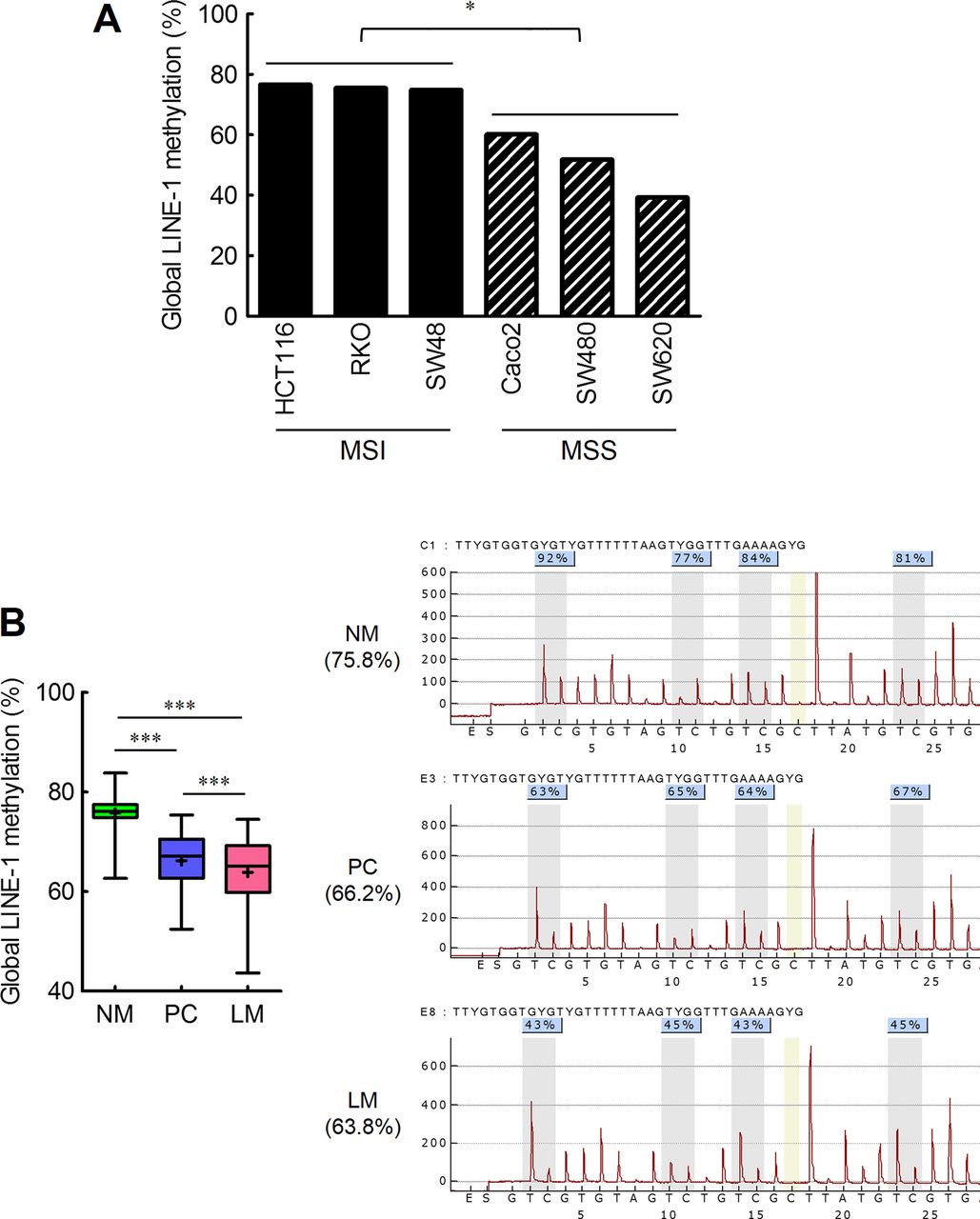
Methylation status of global LINE-1 repeat elements correlates with colorectal cancer (CRC) carcinogenesis as well as liver metastasis. (A) Methylation status of global LINE-1 repeat elements in microsatellite instable (HCT116, RKO and SW48) and microsatellite stable (Caco-2, SW480 and SW620) CRC cell lines (*p<0.05, t test). (B) Methylation status of global LINE-1element in 77 paired normal colonic mucosa (NM), primary CRC tissues (PC), and matched corresponding liver metastasis tissues (LM). (black cross marks represent mean methylation levels; grey vertical boxes in the pyrograms illustrate individual CpG sites analysed; ***p<0.001, t test).
Next, in order to elucidate the contribution of LINE-1 methylation to CRC metastasis, the methylation status of global LINE-1 sequences was measured by quantitative pyrosequencing in matched corresponding NM, PC, and LM tissues (figure 1B and table 1). Compared with NM, both PC (p<0.001) and LM (p<0.001) were significantly hypomethylated at LINE-1 elements. Furthermore, LINE-1 sequences demonstrated significant LINE-1 hypomethylation in LM compared with PC (PC=71.60 (66.2%) vs LM=51.44 (63.8%), mean rank (mean methylation %); p<0.001). Taken together, these data suggest that LINE-1 hypomethylation may play a key role in the development of CRC progression to metastasis.
Global and local LINE-1 methylation in paired NM, PC and LM
LINE-1 hypomethylation correlates with induction of downstream gene transcripts
We wanted to determine the biological consequences of LINE-1 hypomethylation in CRC and liver metastasis. Recent evidence suggests that few genes downstream of LINE-1 elements may be inadvertently reactivated due to LINE-1 hypomethylation, a process that is driven by the antisense promoter for these genes located inside LINE-1 elements.15 ,16 ,26 Using in silico analysis, we discovered three such genes, including MET, RAB3IP and CHRM3, all of which have LINE-1 sequences within their promoter regions (figure 2A, see online supplementary table S3 and figures S4A and S5A). Although MET in linked with CRC metastasis, the molecular mechanisms underlying its activation remain poorly understood.38 ,39 Likewise, both RAB3IP and CHRM3 have been loosely associated with CRC.40 ,41 We, therefore, hypothesised that not just on a global basis, but hypomethylation of local LINE-1 sequences located within the promoter regions of these oncogenes may permit their reactivation and potentially facilitate CRC progression.
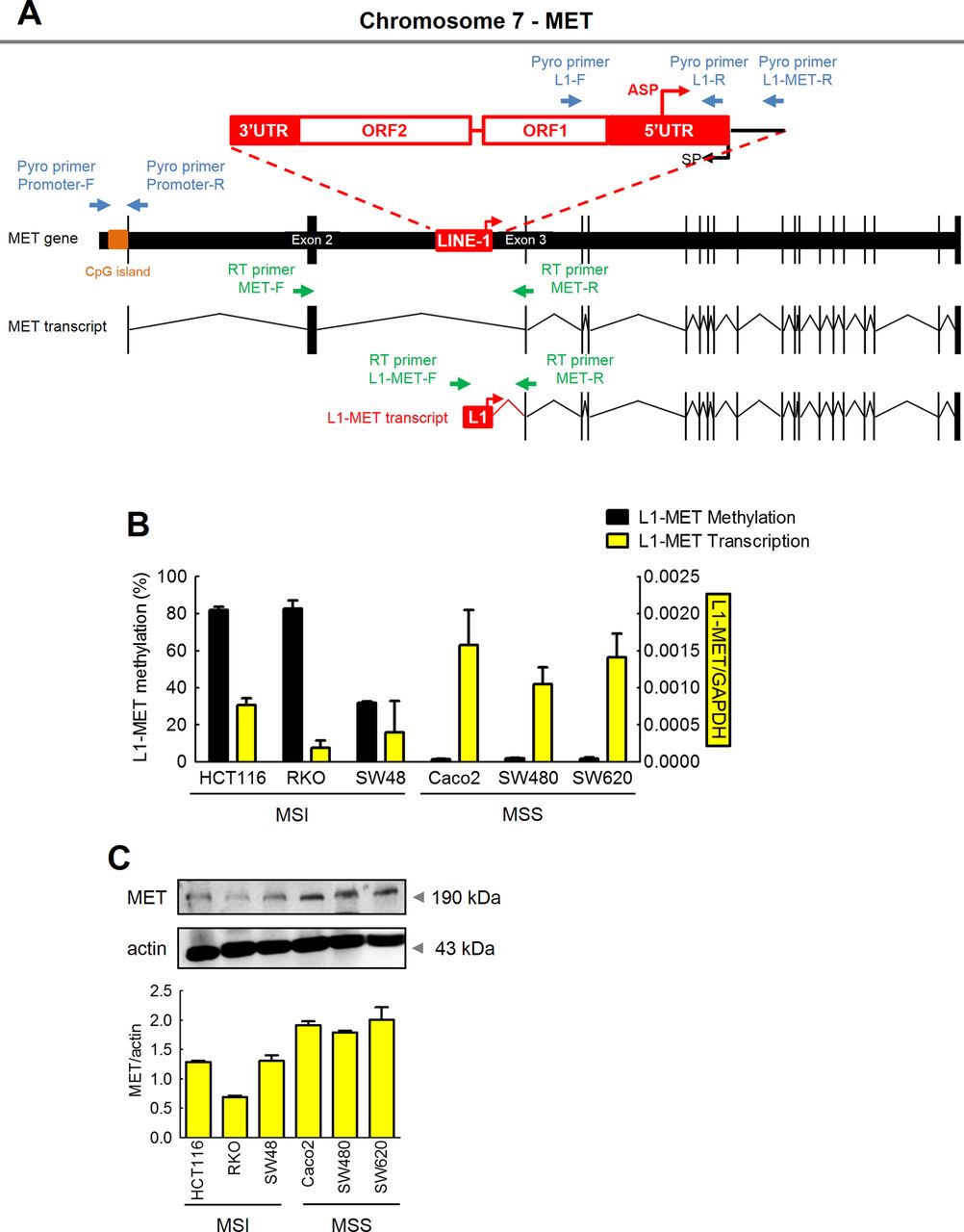
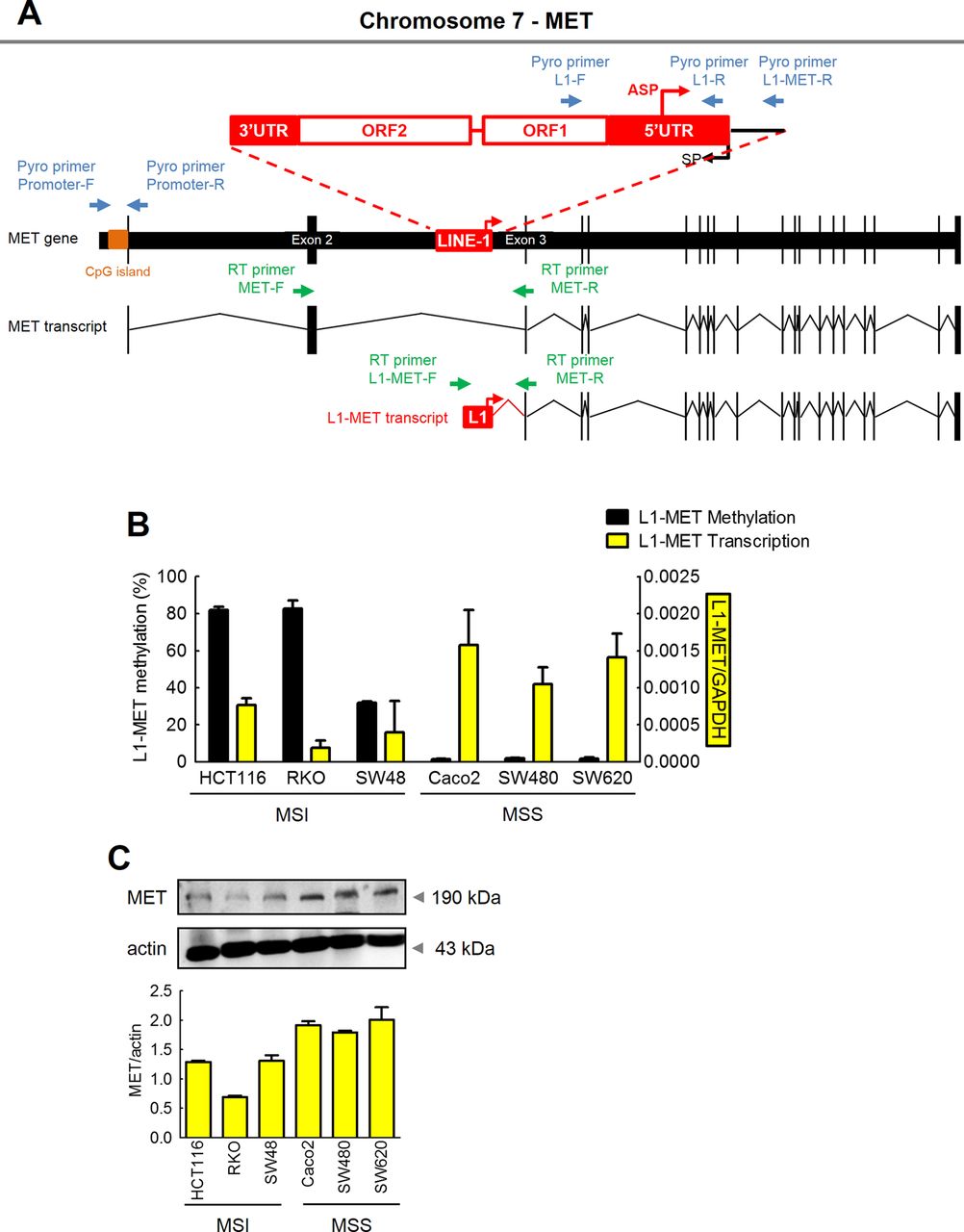
Expression of MET is primarily regulated by hypomethylation of the LINE-1 elements within the host MET gene. (A) Schematic illustration of the local LINE-1 sequence in the MET gene (the red box represents the local LINE-1 element; black boxes represent exons; the red arrow indicates the LINE-1 antisense promoter; blue arrows represent pyrosequencing primers; orange bar indicates CpG island; green arrows represent qRT-PCR primers). (B) Methylation status and transcription status of L1-MET in colorectal cancer (CRC) cell lines was determined by quantitative pyrosequencing analysis and qRT-PCR, respectively. (C) Protein expression (western blotting) of host MET in CRC cell lines.
In order to distinguish between global and local levels of LINE-1 methylation, we designed specific primer sets, in which the forward PCR primer was located inside the LINE-1 promoter, and the reverse primer was located within the intronic region of the host oncogene, allowing measurement of LINE-1 methylation levels that are specific to each gene (figure 2A). Using this approach, we first performed comprehensive analysis of the MET gene. We observed that L1-MET sequences (LINE-1 repeats located within the MET gene intron between exons 2 and 3) were highly methylated in all MSI cell lines (HCT116=82.0%, RKO=82.7%, and SW48=31.8%) compared with MSS cell lines (Caco-2=1.6%, SW480=2.1%, and SW620=1.8%; figure 2B). These results are consistent with the trend observed for global LINE-1 methylation in CRC cell lines (figure 1A), indicating that methylation levels of L1-MET correlate with global hypomethylation in CRC cell lines (figure 1A,B). In order to further understand the functional significance of L1-MET hypomethylation, MSI CRC cell lines with high levels of methylation at L1-MET repeat sequences were treated with the demethylating agent 5-aza-dC. Demethylation of the L1-MET region (HCT116 control=82.0% vs 5-Aza=33.1%; RKO control=82.7% vs 5-Aza=53.6%; SW48 control=31.8% vs 5-Aza=12.9%) resulted in significant hypomethylation of L1-MET as well as activation of L1-MET transcript and MET protein in all MSI cell lines (see online supplementary figure S1A,B). Likewise, L1-RAB3IP and L1-CHRM3 as well as their protein expression were upregulated in CRC cell lines treated with the 5-aza-dC compared with untreated cells, respectively (see online supplementary figure S2A,B).
In addition to methylation status, we also analysed the expression of the LINE-1-induced MET transcript (L1-MET) in CRC cell lines and tissues using highly specific assays. We designed primer sets to distinguish host MET expression and L1-MET expression, in which the forward primers were located either within exon 2 of the MET oncogene or mapped inside the local LINE-1 element, using a common reverse primer located in exon 3 of the MET gene (figure 2A). Demethylation of the L1-MET region induced upregulation of L1-MET transcripts (figure 2B), providing evidence that increased hypomethylation of LINE-1 elements induces activation of downstream gene transcripts.
Increased LINE-1 hypomethylation is associated with overexpression of MET proto-oncogene in CRC cell lines
Previously, overexpression of MET has been observed in various cancers including CRC.38 ,39 Since the MET gene has a CpG island in its promoter region, we determined whether high expression of MET proto-oncogene in CRC may be due to hypomethylation of the host MET promoter region or the L1-MET region.
Unlike L1-MET methylation, negligible levels of host MET promoter methylation were seen in either MSI or MSS CRC cell lines (see online supplementary figure S3A). Next, we determined the relationship between the expression of the MET gene and the methylation status of the L1-MET region. Host MET promoter methylation status was not related with either mRNA or protein expression of the MET gene (see online supplementary figure S3A). By contrast, L1-MET methylation inversely correlated with mRNA expression of L1-MET (figure 2B). Furthermore, L1-MET mRNA expression was consistent with MET protein expression (figure 2B,C). These data provide novel and previously unrecognised insights into the regulatory mechanism underlying the overexpression MET in cancer, and indicate that the high expression of the MET in CRC is primarily driven by hypomethylation of the local LINE-1 sequences residing within the host MET gene.
Hypomethylation of local LINE-1 sequence activates MET proto-oncogene in CRC metastasis
We next tested the hypothesis that LINE-1 hypomethylation would be present in tissue specimens from patients with CRC and LM. As shown in figure 3A, we analysed methylation of L1-MET in 77 matched NM, PC and LM. The PC (66.1%; p<0.001) and LM (49.4%; p<0.001) tissues were significantly hypomethylated compared with NM (91.8%). Moreover, levels of L1-MET methylation were significantly lower in LM compared with PC tissues (p<0.001). By contrast with LI-MET, host MET promoter methylation levels were very low and similar in all NM, PC and LM samples (see online supplementary figure S3B), highlighting the potential significance of LINE-1-specific MET methylation in CRC and metastasis.

Hypomethylation of L1-MET induces transcription of the MET proto-oncogene in colorectal cancer (CRC) metastasis development. (A) Methylation status of L1-MET in normal colonic mucosa (NM), primary CRC tissues (PC), and matched corresponding liver metastasis tissues (LM) (black cross marks represent mean methylation levels; grey vertical boxes in the pyrograms illustrate individual CpG sites analysed; ***p<0.001, t test) (B) Expression status of the L1-MET transcript in PC and matched LM. (**p<0.01, t test) (C) Correlation analysis between L1-MET methylation levels (%) and expression status of L1-MET transcripts (Pearson's correlation coefficient, R; p values, p; red line, linear regression line).
To investigate the functional involvement of LINE-1 hypomethylation in CRC metastasis, expression of LINE-1-induced MET transcription was measured by qRT-PCR in matched corresponding PC and LM (figure 3B and table 1). L1-MET transcription was significantly higher in LM compared with PC (p=0.01). Likewise, the expression pattern of the L1-MET transcript was significantly and inversely correlated with the L1-MET methylation status in CRC tissues (R=−0.44; p<0.0001; figure 3C). On the contrary, host-MET expression did not show a significant difference between matched PC and LM (see online supplementary figure S3C).
In addition to MET transcription analysis, we also determined MET protein expression in matched PC and LM tissues immunohistochemcially (figure 4). The commercially available anti-MET antibodies are unable to discriminate between host-MET and L1-MET protein. Nonetheless, among the 72 pairs of matched PC and LM tissues, 22 (31%) cases showed overexpression of MET protein in LM compared with PC. Forty-four (61%) cases revealed MET overexpression in both PC and LM, whereas only 6 (8%) cases displayed lower expression of MET in LM. These data are of significance as they directly support our observation of upregulation of L1-MET transcription in LM compared with PC.

MET protein is highly expressed in liver metastasis (LM) compared with primary colorectal cancer (CRC) (PC). (A) Representative photomicrographs from a CRC with positive and negative MET protein expression. Among the 72 paired PC and LM tissues, (B) 22 cases showed overexpression of MET in LM, (C) 44 cases revealed comparable expression in PC and LM and (D) 6 cases displayed low expression in LM compared with matched PC.
In order to ascertain whether the consequences of LINE-1 hypomethylation manifest themselves in other LINE-1 associated proto-oncogenes, we analysed the methylation and expression status of two such genes: RAB3IP and CHRM3. We observed that, as was the case with MET, intronic LINE-1 sequences of the RAB3IP and CHRM3 proto-oncogenes, which are upregulated in LM, were significantly hypomethylated in LM compared with PC (RAB3IP, p=0.05; CHRM3, p=0.01; see online supplementary figures S4B,D and S5B,C and table S1). However, host RAB3IP promoter methylation levels were very low in matched PC and LM (see online supplementary figure S4C). These findings have important biological and clinical implications as they suggest that hypomethylation of intronic LINE-1 elements residing within proto-oncogenes can inadvertently result in the activation of various tumour-promoting genes, providing mechanistic insights into the molecular processes that facilitate CRC progression and metastasis development.
Since MET gene amplification in CRC has been previously reported,39 ,42 ,43 we also analysed MET gene copy number variations in CRC cell lines as well as NM, PC and LM tissues. Although MET expression was upregulated in MSS cell lines and 5-aza-dC-treated cell lines, MET gene copy number was not significantly different between MSI and MSS cell lines, as well as between CRC cell lines treated with the 5-aza-dC and non-treated cells (see online supplementary figure S6A). Although MET gene copy number was significantly amplified in the PC and LM tissues compared with NM tissues (see online supplementary figure S6B), no significant differences were observed between matched PC and LM tissues.
5-hmc associates with hypomethylation of LINE-1 in CRC
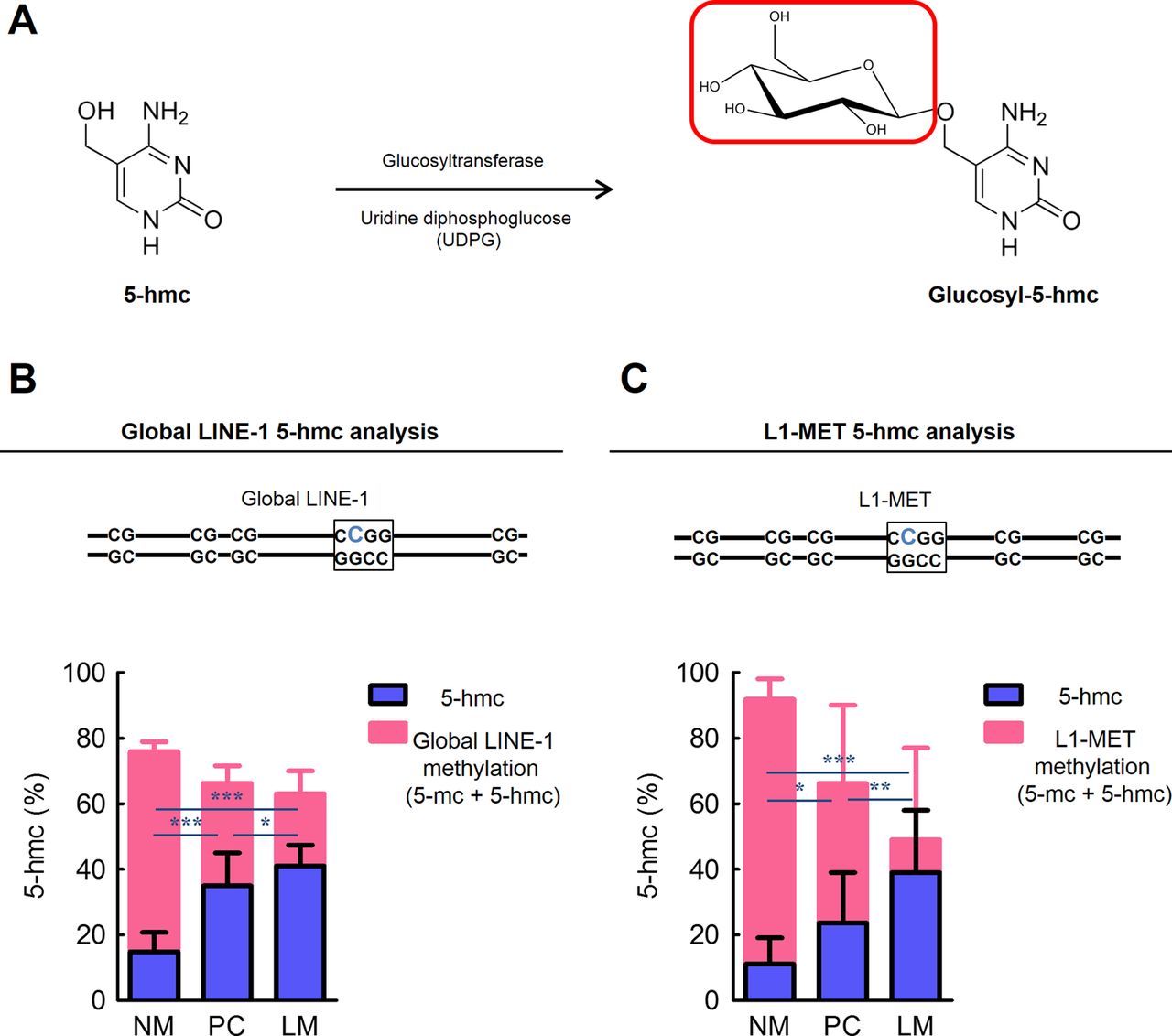
Recently, it has been suggested that the presence of 5-hmc precedes DNA demethylation.33–35 Since conventional methylation analysis methods, such as bisulfite modification-based methods and restriction enzymatic approaches are unable to discriminate between 5-mc and 5-hmc,33 ,44 we applied a novel analysis that involves the generation of glucosyl-5-hmc for the detection of locus-specific 5-hmc (figure 5A).

The presence of 5-hydroxymethylcytosine (5-hmc) is related to demethylation of LINE-1 sequence in colorectal cancer (CRC) metastasis. (A) Glucosyltransferase catalyses the transfer of a glucose moiety from uridine diphosphoglucose onto the 5-hmc. (the red box indicates the transferred glucose moiety) (B) The quantity of 5-hmc at a CpG locus in global LINE-1 and (C) in local L1-MET in normal colonic mucosa (NM), primary CRC tissues (PC), and matched liver metastasis tissues (LM). (the blue bold ‘C’ character indicates the CpG locus analysed; *p<0.05, **p<0.01, ***p<0.001, t test).
We measured 5-hmc levels at the CpG locus located in global LINE-1 or local L1-MET by qRT-PCR in matched NM, PC and LM (figure 5B,C). By contrast with global LINE-1 methylation, the 5-hmc content in global LINE-1 elements in PC (p<0.001) and LM (p<0.001) tissues were significantly higher compared with NM. Moreover, the 5-hmc levels in global LINE-1 elements in LM were significantly elevated compared with PC (p<0.05). Similarly, the amount of 5-hmc in local L1-MET was significantly increased in PC (p<0.05) and LM (p<0.001) compared with NM, respectively. Additionally, LM showed significantly higher amounts of 5-hmc in local L1-MET compared with PC (p<0.01). Taken together, these findings suggest that increased amounts of 5-hmc are potentially involved in active LINE-1 hypomethylation in colorectal carcinogenesis and progression.
Activation of MET oncogene triggered by aberrant LINE-1 hypomethylation associates with more aggressive progression of CRC metastasis
To determine associations between LINE-1 methylation alterations and time to metastasis, we analysed global LINE-1 methylation, L1-MET methylation and L1-MET expression in PC tissues with metachronous liver metastasis (M-CRC) and synchronous liver metastasis (S-CRC; see online supplementary figure S7). Both global LINE-1 and L1-MET sequences were significantly hypomethylated in S-CRC compared with M-CRC which was further supported by significant upregulation of L1-MET transcripts in S-CRC compared with M-CRC.
We next explored the relationship between MSI status, and global as well as LI-MET hypomethylation in CRC tissues. Although our cohort included only 2 MSI-positive CRCs prohibiting a meaningful statistical analysis (see online supplementary table S1), these tumours showed increased global LINE-1 methylation as well as L1-MET methylation together with corresponding suppression of L1-MET transcripts (see online supplementary figure S8A) compared with MSS CRCs. With regards to the relationship between time to liver metastasis recurrence and alterations in LINE-1 hypomethylation, we analysed its status in M-CRC tissues (see online supplementary figure S8B). Although there was a solitary case of M-CRC with MSI, this tumour did exhibit prolonged disease-free survival and higher global LINE-1 methylation and L1-MET methylation as well as lower expression of L1-MET expression compared with MSS M-CRC patients. These data suggest that activation of MET oncogene triggered by aberrant LINE-1 hypomethylation in MSS CRC could be the reason for poor prognosis and high frequency of metastasis typically observed in MSS CRCs.
Discussion
Although a growing body of scientific evidence indicates the involvement of global hypomethylation of LINE-1 retrotransposable elements in carcinogenesis, the consequences of these events in driving malignant progression remain poorly understood.15–25 In the context of CRC, the major cause of morbidity and mortality is advanced disease and distant metastasis. In this study, we provide the first evidence for increased LINE-1 hypomethylation in CRC metastasis, and systematically uncover its relationship with hypomethylation of specific LINE-1 sequences that occur in the context of three proto-oncogenes, for example, MET, RAB3IP and CHRM3 in primary CRC and liver metastasis. More specifically, our present study demonstrates: (1) that global LINE-1 sequences are frequently hypomethylated in primary CRC, and significantly more prevalent in liver metastases in CRC patients; (2) that hypomethylation of specific LINE-1 elements located upstream of certain proto-oncogenes permit transcriptional activation of three such oncogenes (ie, MET, RAB3IP and CHRM3) which occurs more frequently in liver metastasis versus primary CRC; (3) that higher expression of MET protein is observed in liver metastasis compared with primary CRC which specifically correlates with higher levels of L1-MET transcription; (4) that increased amounts of 5-hmc correlate with hypomethylation of LINE-1 sequences, suggesting that the presence of 5-hmc plays a causal role in aberrant LINE-1 hypomethylation in human cancer.
Although similar to this study, others have previously reported the existence of global LINE-1 methylation in CRC,23–25 none of these studies has investigated the functional role of LINE-1 hypomethylation in the development of metastasis in human CRC. This is the first study which analyses the methylation status of LINE-1 repeat elements in normal colonic mucosa, primary cancer tissues (CRC, in this instance), and matching metastatic tissues (liver, in this case). Our data reveal that global LINE-1 hypomethylation significantly increases during CRC progression, and this stepwise process follows the following order: normal colonic mucosa, primary CRC and liver metastasis. We also noted that LINE-1 sequences were significantly hypomethylated in SW620 (derived from a lymph node metastasis from the same patient as SW480) compared with SW480 cells, lending further credence to our hypothesis for the involvement of LINE-1 hypomethylation during CRC progression. Additionally, it has been shown that hypomethylation of LINE-1 sequences is absent in the non-cancerous primary liver tissues, and even in tissues with liver cirrhosis,22 suggesting LINE-1 hypomethylation is an identifying mark of metastasised CRC in liver. Taken together, these results not only strengthen the importance of LINE-1 hypomethylation in CRC development, but also highlight their novel potential role in CRC metastasis development.
Another novel aspect of this study was that we successfully elucidated the oncogenic role of LINE-1 hypomethylation in CRC metastasis. Although few previous studies have shown the activation of MET gene transcription by upstream LINE-1 antisense promoters in chronic myeloid leukaemia,16 bladder cancer15 and CRC cell lines,45 ours is the first study that systematically interrogated hypomethylation of intronic-LINE-1 elements and demonstrates transcriptional activation of three such proto-oncogenes (MET, RAB3IP and CHRM3) mediated by a LINE-1 internal antisense promoter (ASP) in cancer metastasis using primary CRC and matching liver metastasis. Because we were able to design unique assays to determine the expression of LINE-1 sequences in the regulatory regions of various oncogenes including MET, we were able to successfully delineate the differences in expression of the host MET gene transcript versus the LINE-1 ASP-induced transcript. More importantly, we first demonstrated that the changes in MET protein expression status not only correlated with the LINE-1 ASP-induced MET transcript, but also correlated inversely with LINE-1 ASP methylation. Interestingly, neither MET host promoter methylation nor the host MET transcript correlated with MET protein expression; rather it was regulated by the methylation status of the LINE-1 element. With regards to the activation mechanism of MET expression in CRC, MET gene amplification has been reported previously,39 ,42 ,43 but there has been a lack of consensus on copy number variations and disease pathogenesis due to differences in analytical methods and the MET gene locus interrogated in these previous articles. In our study, MET gene copy numbers were significantly higher in primary CRC and liver metastasis compared with normal mucosa, while no significant differences were observed between matched primary CRC and liver metastasis. Our findings provide a novel mechanistic insight that was not recognised in previous studies,42 ,46 and suggests that while global MET gene amplification is associated with the development of primary colorectal carcinogenesis, CRC metastasis may be primarily associated with the activation of specific L1-MET transcripts that are triggered by LINE-1 hypomethylation.
In terms of the clinical relevance of LINE-1 element variation, LINE-1 hypomethylation-induced MET activation in cancer provides important evidence for aberrant LINE-1 methylation in MSS CRC which is associated with poor prognosis and a high frequency of metastasis.23 We noted that both global LINE-1 and L1-MET sequences were significantly hypomethylated in S-CRC compared with M-CRC. These data were further supported by significantly upregulated expression of L1-MET transcripts in S-CRC vis-a-vis M-CRC. Not surprisingly, we discovered that only two cases were MSI-positive, as MSI neoplasms are less often metastatic. Both MSI CRCs showed increased global LINE-1 methylation, as well as L1-MET methylation, together with corresponding suppression of L1-MET transcripts. Furthermore, although there was a single M-CRC case with MSI, this patient exhibited prolonged disease-free survival and higher global LINE-1 methylation and L1-MET methylation, as well as lower expression of L1-MET expression compared with MSS M-CRC patients.
In addition to MET, we also showed that LINE-1 hypomethylation leads to the activation of two other oncogenes, RAB3IP and CHRM3. Although the biological function of RAB3IP related to CRC metastasis is not well established, it has been shown that RAB3IP protein shuttles cell-cycle-related factors between the nucleus and the cytoplasm,41 as well as being responsible for alternative splicing of the RAB3IP gene transcripts in human cancers.47 ,48 Similarly, CHRM3 is frequently overexpressed in CRC cell lines and tissues, and participates in enhanced cell proliferation, survival, migration and invasion, in part, by a COX-2-mediated cellular mechanism.40 ,49 ,50 Taken together, our results provide novel mechanistic insights into the molecular mechanism underlying the activation of the MET proto-oncogene, and highlight the functional importance of LINE-1 ASP hypomethylation in mediating the induction of this key proto-oncogene in CRC.
In light of the previous reports that 5-hmc may play an important role in the regulation of DNA demethylation, we analysed 5-hmc content that is specifically associated with LINE-1 sequences in human CRC tissues. This is of consequence, because 5-hmc may reduce the binding affinity of 5-mc-binding proteins (MBPs), such as MBD1, MBD2, MBD4 and MeCP2.35 The conversion of 5-mc to 5-hmc abrogates DNMT1 activity that can result in the failure of maintenance methylation during cell division.34 Also, 5-hmc may interact with specific chromatin-modifying partners, thus altering chromatin structure and DNA methylation.33 Since previous reports have not found a direct correlation between global 5-mc level and LINE-1 methylation,51 in this study, we measured LINE-1 sequence-specific 5-hmc content instead of global 5-hmc levels. We found that an increase in 5-hmc is related to LINE-1 hypomethylation in CRC tissues. Unlike in tissues, we observed only a modest level of 5-hmc in CRC cell lines (data not shown) which is consistent with recent reports. Although the detailed mechanisms should be further investigated, we observed for the first time that a change in the amount of 5-hmc correlated with LINE-1 hypomethylation in human cancer tissues.
In view of the comprehensive data presented in this study, we propose a consolidated model that systematically summarises the potential role of LINE-1 hypomethylation in CRC progression (figure 6). According to this model, DNA hypomethylation of CpG sites is best characterised by the conversion of 5-mc (5-methyl cytosine) to 5-c (5-cytosine) in a sequential manner. During the first step, 5-mc undergoes a hydroxylation step to generate 5-hmc (5-hydroxy methyl cytosine) which prevents binding of DNA methylation-related enzymes, such as DNMT1 and MBPs, that drive DNA hypermethylation, leading to reduced DNA methylation and a simultaneous accumulation of 5-c (5-cytosine). During CRC metastasis, 5-mc to 5-c conversion at CpG dinucleotides occurs in a genome-wide manner including LINE-1 elements. However, when such events occur in the LINE-1 sequences, it results in their demethylation, and also permits activation of the ‘constitutionally methylated’ ASP, facilitating the transcription of various downstream proto-oncogenes. Herein, we provide evidence that MET is one such gene, whose expression is controlled by the methylation status of a LINE-1 transposable element, and may play a role in accelerating the development of CRC metastasis.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
A consolidated model for the activation mechanism of the MET proto-oncogene by LINE-1 hypomethylation in colorectal cancer (CRC) metastasis. In this model, hypomethylation of CpG sites is characterised by enhanced conversion of 5-mc (5-methylcytosine) to 5-c (5-cytosine), with an intermediary generation of 5-hmc (5-hydroylated methylcytosine). Enhanced CpG demethylation with consequent accumulation of 5-hmc prevents the binding of various DNA methyltransferases (DNMTs) and methyl binding proteins (MBPs) to CpG sites, triggering a cascade of events that favour DNA hypomethylation. Occurrence of DNA hypomethylation events in the context of LINE-1 elements can potentially permit activation of certain oncogenes, including the MET proto-oncogene which may result in accelerated CRC metastasis.
In conclusion, our data highlights the pivotal role of LINE-1 hypomethylation in CRC metastasis development. In this study, we provide mechanistic evidence that links the LINE-1 hypomethylation with the induction of proto-oncogenes in CRC metastasis. Moreover, we provide unique evidence that the amount of 5-hmc correlated with LINE-1 sequence demethylation in cancer tissues. These results underscore the potential diagnostic and prognostic value of LINE-1 hypomethylation as a biomarker for metastatic cancer.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online figure 1
- Data supplement 2 - Online figure 2
- Data supplement 3 - Online figure 3
- Data supplement 4 - Online figure 4
- Data supplement 5 - Online figure 5
- Data supplement 6 - Online figure 6
- Data supplement 7 - Online figure 7
- Data supplement 8 - Online figure 8
- Data supplement 9 - Online legends
Footnotes
-
Contributors Study concept and design (KH and AG); provision of samples (PC, JM-R, EB and JF); acquisition of data (KH); analysis and interpretation of data (KH and AG); statistical analysis (KH); drafting of the manuscript (KH, CRB and AG).
-
Funding The present work was supported by grants R01 CA72851 and CA129286 from the National Cancer Institute, National Institutes of Health, and funds from the Baylor Research Institute to CRB and AG.
-
Competing interests None.
-
Patient consent Obtained.
-
Ethics approval Institutional Review Board.
-
Provenance and peer review Not commissioned; externally peer reviewed.