

Abstract
Brainstem nuclei have well established roles in generating nonlearned rhythmic behaviors or as output pathways for more complex, forebrain-generated behaviors. However, the role of the brainstem in providing information to the forebrain that is used to initiate or assist in the control of complex behaviors is poorly understood. In this study, we used electrical microstimulation in select nuclei of the avian song system combined with recordings of acoustic and respiratory output to examine how forebrain and brainstem nuclei interact in the generation of learned vocal motor sequences. We found that brief stimulation in the forebrain nuclei HVC (used as a proper name) and RA (robust nucleus of the arcopallium) caused a short-latency truncation of ongoing song syllables, which ultimately led to a cessation of the ongoing motor sequence. Stimulation within the brainstem inspiratory-related nucleus paraambigualis, which receives input from RA and projects back to HVC via the thalamus, caused syllable truncations and interruptions similar to those observed in HVC and RA. In contrast, stimulation in the tracheosyringal portion of the hypoglossal nucleus, which innervates the syrinx (the avian vocal organ) but possesses no known projections back into the song system, did not cause any significant changes in the song motor pattern. These findings suggest that perturbation of premotor activity in any nucleus within the recurrent song system motor network will disrupt the ongoing song motor sequence. Given the anatomical organization of this network, our results are consistent with a model in which the brainstem respiratory nuclei form an integral part of the song motor programming network by providing timing signals to song control nuclei in the forebrain.
Introduction
Much is known about the role that brainstem nuclei play in generating relatively simple or nonlearned rhythmic behaviors (Cohen et al., 1996; Sparks, 2002; Feldman et al., 2003) or as output pathways for more complex behaviors that are ultimately under the control of the forebrain (Jürgens, 2002; Krauzlis, 2004). Much less is known about how brainstem centers provide instruction to the forebrain to initiate or assist in the control of complex behaviors. Although the presence of recurrent pathways from the brainstem into the forebrain suggests that brainstem nuclei can perform these roles (Striedter and Vu, 1998; Jürgens, 2002; Sommer and Wurtz, 2004), the potential contributions to complex motor program generation made by these nuclei remain essentially unexplored.
The avian song system is one example of a system in which both forebrain and brainstem centers interact to produce complex, stereotyped, learned behavior. In the song system, several telencephalic nuclei are critical to song behavior, and lesions of these, such as the forebrain nucleus HVC (used as a proper name), or of one of its efferent targets, the robust nucleus of the arcopallium (RA), disrupt or prevent song production (Nottebohm et al., 1976; Simpson and Vicario, 1990; Halle et al., 2003). Electrical microstimulation of these structures also affects song production (Vicario and Simpson, 1995; Fee et al., 2004) and, when applied to HVC in particular, stops or restarts ongoing song in both auditorily intact and experimentally deafened birds, implying that the stimulation effects cannot be attributed to auditory feedback (Vu et al., 1994, 1998). The descending projections of RA terminate in brainstem nuclei that are necessary for song production (Vicario, 1991), namely the tracheosyringal portion of the hypoglossal nucleus (nXIIts) that controls muscles of the syrinx, the avian vocal organ, and medullary premotor nuclei that, via their spinal projections, modulate the activity of inspiratory and expiratory motor neurons (Nottebohm et al., 1976, 1982; Wild, 1993a,b; Reinke and Wild, 1998) (Fig. 1). These same respiratory premotor nuclei also have ascending projections, and one of these nuclei, the inspiratory-related nucleus paraambigualis (PAm), sends bilateral projections specifically to nucleus uvaeformis (Uva) in the posterodorsal thalamus, which is a major source of direct and indirect ipsilateral inputs to HVC (Nottebohm et al., 1982; Reinke and Wild, 1998; Striedter and Vu, 1998; Coleman and Vu, 2005). These ascending projections could provide important song-related information or feedback to forebrain song nuclei from the brainstem.
Premotor activity within HVC is synchronized between hemispheres during song production (Schmidt, 2003), despite the absence of commissural connections between these two nuclei or any other forebrain song control nuclei (Fig. 1). These observations suggest that the bilaterally projecting brainstem respiratory centers may provide synchronizing input into the forebrain song system. This hypothesis contrasts with previously proposed models of song production in which HVC has been hypothesized to serve as a more autonomous motor program generator for song.
In the present study, we used electrical microstimulation combined with recordings of acoustic output and air sac pressure to study the relationship between specific forebrain and brainstem nuclei and their contribution to song production. Our results suggest that, rather than simply following instructions from the forebrain, brainstem respiratory nuclei play a critical role in the generation of the song motor patterns. The results suggest that the recurrent brainstem pathways leading back into the forebrain play an important role in guiding and coordinating HVC activity in its control of song production.
Materials and Methods
Animals. Adult male zebra finches (Taeniopygia guttata) ranging from 120 to 500 d of age were obtained from our breeding colony and from a local supplier. Birds were housed under constant 12 h light/dark conditions and given food and water ad libitum. All procedures described here were approved by an institutional animal care and use committee at the University of Pennsylvania.
Chronic electrode implants. Methods for implanting chronic stimulating electrodes have been described previously (Vu et al., 1998). Briefly, adult male zebra finches were anesthetized with 0.7–0.8 ml of ketamine/xylazine (40 mg/kg ketamine and 8 mg/kg xylazine; Phoenix Pharmaceuticals, Mountain View, CA). HVC, RA, PAm, and nXIIts were identified using stereotaxic coordinates combined with electrophysiological recordings to search for characteristic neural activity patterns. For HVC and RA, this consisted of spontaneous activity that was considerably higher than surrounding areas and frequently appeared as arrhythmic bursts. PAm was identified at previously determined stereotaxic coordinates (Reinke and Wild, 1998) in conjunction with recordings of a strong respiratory rhythm in phase with inspiration, as determined by air sac recordings and correlated signs of thoracic expansion. In one bird, neural recordings were also made through the stimulation electrode during singing, and the presence of inspiratory-related activity was verified. Electrode placement for another bird was verified by frozen-section histology. The location of nXIIts was identified by stereotaxic coordinates and the presence of a strong respiratory rhythm. Two birds received electrode implants in one nXIIts. A third bird received electrodes in both right and left nXIIts. All nXIIts electrode implant locations were verified by frozen section histology. Of these four electrodes, one was on the rostral, ventral, and lateral edge of nXIIts, one was centrally located, and two (from the double implant) were each located in the medial portion of the nucleus.
Electrodes were constructed from either 25 μm Formvar-insulated nichrome wire (HVC and RA; AM Systems, Carlsborg, WA) or from 100 μm Teflon-coated tungsten electrodes (RA, nXIIts, and PAm; Fredrick Haer Company, Bowdoinham, ME). Impedance of these electrodes ranged from 200 kΩ to 1 MΩ. These were attached to three- or four-pin microconnectors (Omnetics, Minneapolis, MN). After correct placement of each electrode, the exposed ends and the connector were cemented to the skull using dental cement (Dentsply International, Milford, DE). Birds were allowed to recover for ≥2 d before performing stimulation.

Diagram of the avian song system emphasizing its bilateral organization and the bilateral projections from the brainstem to the forebrain. The portion of the song system that is thought to be involved in song pattern generation is highlighted in gray. This loop consists of the forebrain nucleus HVC, the dRA, the vocal-respiratory network (highlighted in darker gray), and Uva. The vocal-respiratory network is made up of RAm, PAm, and DM (dorsomedial nucleus of the intercollicular complex). The thalamic nucleus DMP and MMAN have been suggested to play an indirect role in motor production (Foster and Bottjer, 2001; Coleman and Vu, 2005) (for review, see Schmidt et al., 2004). Although NIf receives inputs from Uva and sends a strong projection to HVC, it does not appear to be directly involved in song motor patterning in the zebra finch (Cardin et al., 2005) (but see Hosino and Okanoya, 2000). The connection between vRA and nXIIts serves as an output pathway to the syrinx rather than as part of the song pattern generator. INSP and EXP represent, respectively, the inspiratory and expiratory motor neurons. The circuit known as the anterior pathway, which is made up of Area X, DLM, and LMAN, is not necessary for song production but plays an important role in song learning and maintenance. Numbers indicate sites at which stimulation was performed. The three parallel lines between hemispheres illustrate the lack of commissural connections between forebrain song control nuclei. Many of the nuclei in the vocal motor system (DMP, RAm, PAm, and DM), however, project directly or indirectly to vocal motor nuclei in the contralateral hemisphere. Although these projections are bilateral, we only illustrate projections from the left to the right hemisphere for simplicity. Nuclei receiving contralateral inputs are highlighted in dark gray. The anatomical connections shown in this diagram represent the major projections in the song system and have been compiled from a number of different sources (Stokes et al., 1974; Nottebohm et al., 1982; Vicario, 1991; Vates et al., 1997; Reinke and Wild, 1998; Striedter and Vu, 1998; Sturdy et al., 2003; Wild, 2004a,b). Weak projections have been left out. DLM, Medial part of the dorsolateral thalamic nucleus; DMP, dorsomedial posterior nucleus of the thalamus; LMAN, lateral magnocellular nucleus of the anterior nidopallium; MMAN, medial magnocellular nucleus of the anterior nidopallium; Area X, Area X of the medial striatum; NIf, nucleus interfacialis of the nidopallium.
Electrical microstimulation. Animals were placed in a sound-attenuating recording chamber (Industrial Acoustics Company, Bronx, NY) and attached to a stimulator (AM Systems) via a wire tether and a mercury commutator, allowing the birds general freedom of movement. After a period of adjustment, the implanted finches sang readily to females that were placed in an adjacent cage. Monopolar stimulation was triggered during singing automatically by computer, using a modified version of the Sound Analysis Live software (Tchernichovski et al., 2000), which also recorded song output and the time of stimulation. Sound was recorded using one to three omni directional microphones (Earthworks, Milford, NH), and each sound channel was sampled at 44,100 Hz. The distance between the bird and the microphone was never more than 30 cm, ensuring that the delay for sound to reach the microphones was always <1 ms. This delay has not been subtracted from measurements of acoustic latency (see below).
Stimulation was triggered by acoustic features, including overall amplitude, power at different frequencies, and Wiener entropy. Stimulation was delivered to only one of the implanted electrodes at a time and was usually delivered once per song with a randomized delay. Unless otherwise noted, stimulation consisted of five 400 μs biphasic pulses delivered over a duration of 12 ms. Stimulus intensities ranged from 15 to 60 μA. These short stimulus durations were not sufficient to evoke vocalizations; however, longer durations (100 ms or more) in HVC and PAm sometimes produced brief, unstructured vocalizations.
Analysis of the effect of stimulation on song was performed by visual inspection of amplitude waveforms and song spectrograms. We used standard nomenclature for defining zebra finch song elements. The smallest vocal unit at least 30 ms long and separated by silent intervals of at least 10 ms is referred to here as a “syllable” (Williams, 1990). Syllables were further subdivided into notes, which were delineated by abrupt changes in acoustic structure. Multiple syllables in a stereotyped order were defined as a motif, with multiple sequential motifs comprising a complete song. Songs from all stimulation intensities were randomly intermixed and scored blind. As part of this randomized set, a collection of songs with no stimulation was included, and, for each of these songs, a random stimulation time (measured from the beginning of the first syllable of the song) was selected. Each stimulation time for this control group was randomly drawn from the distribution of actual stimulation times.
Stimulation effects were divided into two general categories: syllable-level effects and song-level effects.
At the syllable level, two classes of effects were defined: (1) syllable distortion, in which the length of the stimulated syllable was at least 80% of the mean length for that syllable, but the amplitude waveform or spectrogram judged by a blind observer was different from the unstimulated equivalent syllable; and (2) syllable truncation, in which the length of the stimulated syllable was <80% of the mean length for that syllable.
At the song level, two classes of effects were defined: (1) stopping of the song, in which stimulation caused a cessation of vocalizations for at least 500 ms after the stimulated syllable; and (2) restarting of the motif, in which stimulation caused production of a new motif within 500 ms of the stimulated syllable. This new motif was sometimes preceded by introductory notes, a call, or both.
Each song was scored first for syllable truncation, and, if no truncation occurred, songs were examined for evidence of syllable distortion. Therefore, truncation and distortion were mutually exclusive. Regardless of the presence of syllable-level effects, each song was then scored for song-level effects. These were categorized as song stops, motif restarts, or no effect. Song-level effects were, therefore, mutually exclusive. After all songs had been scored, the percentage of songs with each effect was calculated. To determine the significance of the number of song-level and syllable-level effects seen with different treatments (intensity or location), the proportion of effects observed in songs that received stimulation was compared with the proportion of effects scored in control songs (0 μA, no stimulation data). The number of scored songs with syllable-level or song-level effects for a given bird at a given stimulus intensity was compared with the control set of songs for that bird using a heterogeneity G test (Sokal and Rohlf, 2001). A heterogeneity G test is similar to a χ2 test, but it additionally accounts for variability between subjects (birds) when determining significance. Because G test scores are cumulative, we were able to calculate scores for each bird using its own control set of songs and compile these into a pooled G score for each group.
For some stimulation sites, we measured syllable truncation latency as a metric for how quickly stimulation affected song production (for details, see Results). Latency was measured as the duration from the first stimulation pulse to the time that sound returned back to baseline amplitude levels. Multiple truncation times were collected for each bird tested. Significance between groups of birds was measured using a nested ANOVA (NANOVA) (two level with unequal sample size). We attempted to measure the latency between stimulus onset and the time the acoustic amplitude envelope first deviated from the mean envelope for that syllable. The gradual time course of this deviation combined with the high variability of the acoustic envelope, however, prevented reliable estimates.
Air sac pressure recording. Air sac pressure recording was performed as described previously (Franz and Goller, 2002). Briefly, under pentobarbital anesthesia (43 mg/kg), a thin pressure cannula was inserted into the abdominal air sac on the left or right side just below the lowest rib. This cannula was connected to a small pressure transducer (FHM-02PGR-02; Fujikura, Tokyo, Japan) that each bird wore on its back. The transducer was powered by a custom-built power source (provided by Brent Cooper, University of Utah, Salt Lake City, UT), and was connected via a wire tether to a four-channel amplifier (model 440; Brownlee, San Jose, CA). In most cases, the signal was bandpass filtered between 0.1 and 1000 Hz. The signal was acquired through a PCI-MIO-16XE-10 data acquisition device (National Instruments, Austin, TX) using the Multi-Neuron Observer software program written in Labview (National Instruments) by A. Leonardo (California Institute of Technology, Pasadena, CA). To generate average pressure traces for particular motif segments, pressure traces from multiple renditions of the segment were manually aligned with one another so as to maximize the temporal overlap of all traces. The mean and SDs of these traces were then calculated at each time point using custom scripts written with Matlab (MathWorks, Natick, MA). The latency to deviation from the normal song respiratory pattern was determined by manually aligning the respiratory trace for stimulated trials with the average control trace, again attempting to maximize overlap of the traces in the period before stimulus onset. The time of air sac pressure change was defined as the time when the stimulation trace deviated from the control trace by more than 2 SDs. The manual alignment of stimulation-altered traces with control traces was repeated for every instance of individual stimulation.
Because the electrodes and stimulation techniques used here were nearly identical to those used by Vu et al. (1998), it was possible to make meaningful comparisons between the latency to suppression of neural activity reported in that study and the air sac pressure change latency reported in the present study (see Fig. 9). These comparisons were made using Student's t test.
A similar method was used to measure air sac pressure changes resulting from stimulation during quiet respiration. Multiple air sac pressure traces were aligned with stimulation onset and averaged. To determine the time of deviation from baseline, pressure values for a 50 ms period before stimulation were averaged, and a value of 2 SDs above this mean was used as a threshold. Calculation of baseline respiratory amplitude range was made by averaging the absolute differences between minimum and maximum air sac pressure values during randomly selected respiratory cycles.
Histology. Birds were deeply anesthetized with 0.1 ml of 50 mg/ml Nembutal (Abbott Laboratories, Abbott Park, IL) and perfused with 0.9% saline and 4% paraformaldehyde. Brains were cryoprotected in 30% sucrose and sectioned at 40 μm on a freezing microtome. Sections were cut parasagittally for verification of forebrain electrode placement, and coronal sections were used for the identification of brainstem electrode placement. All sections were stained with cresyl violet.
Results
This study consists of three parts. In the first experiment, we stimulated HVC during song and quantified stimulation effects on syllable production. We quantified the effect of stimulation on both acoustic output and the respiratory pattern. In the second experiment, we stimulated different regions of RA to examine the differential contribution of these regions to song patterning. Finally, we stimulated in brainstem areas that serve as output structures for RA to test the role of the respiratory-vocal network in song motor programming.
Temporary disruption of HVC premotor activity causes truncation of ongoing syllables
To test the effects of stimulation in HVC on individual syllables and whole motifs, we developed an automated system to deliver brief stimulation pulses during song production (for details of stimulation parameters, see Materials and Methods). Using this system, we were able to analyze the effect of stimulation on song for a larger dataset (n = 3098 songs in 4 birds) and in a much more detailed way than in previous studies (Vu et al., 1994, 1998). In agreement with these previous studies, we found that HVC stimulation disrupted both acoustic and temporal features of the song (Figs. 2, 3).

Effects on song from stimulation in HVC. A, Syllable-level effects. The same syllable is shown in its normal unstimulated state (left), truncated by a brief stimulus applied to HVC (center), and distorted by a similar intensity stimulus (right). The arrow indicates the time of stimulation in each case. B, Song-level effects. The top most sonogram is an example of a normal song. In the example below it, microstimulation caused the ongoing motif to stop prematurely and then restart with a new motif (motif restart) after producing two introductory notes (i). In the bottom example, microstimulation caused the ongoing motif to stop prematurely (song stop), with no subsequent song production. In both of these examples, song stops and restarts are accompanied by a truncation of the ongoing syllable. Alphabetical letters symbolize the different syllables in each motif. A* indicates a truncated version of syllable A.
To assess the overall effect of unilateral HVC stimulation during singing, data were compiled for all birds and divided into two categories. The first category, shown in Figure 2A, consisted of syllable-level effects. Effects in this category reflect those songs in which HVC was stimulated and either the ongoing syllable was truncated prematurely (“truncate”) or the amplitude and/or spectral content of the ongoing syllable were changed (“distort”). Songs were scored as containing either syllable truncation or distortion but not both (for a detailed definition of these categories, see Materials and Methods). Over half of the songs in which 15–60 μA stimulation was delivered (Fig. 3A) showed syllable-level effects (distortions or truncations, 51.5%; n = 1409 of 2735 songs in 4 birds). To make an overall assessment of stimulation effects, we calculated the percentage of all songs for all birds that showed syllable-level effects at each stimulus intensity, ranging from 15 to 60 μA. These percentages were compared with a control set of songs in which no stimulus was delivered and which were nonetheless scored for syllable effects (see Materials and Methods).

Quantification of syllable and song-level effects after stimulation in HVC. A, Total number of songs showing different effects from HVC stimulation, of a total of 2735 stimulated songs in four birds. These values represent stimulation at intensities ranging from 15 to 60μA. Syllable-level effects are shown on the left, comprising either truncation of the ongoing syllable (truncate, light gray) or acoustic and/or spectral distortion of the stimulated syllable (distort, dark gray). Syllables in which stimulation had no effect are not shown. These two effects are mutually exclusive. Song-level effects are shown on the right and comprise song stopping (stop, light gray) or stopping followed by restarts of a new motif (restart, dark gray). These two effects are also mutually exclusive. B, Syllable-level effects resulting from HVC stimulation at different current intensities. C, Song-level effects resulting from HVC stimulation at different current intensities. All categories in B and C are compared with percentages collected for a control set of songs with no stimulation (CONT). Significance levels were measured using a G test comparing each stimulus intensity category with the control category for each bird. *p < 0.001. D, Relationship between syllable truncation and song-level effects. Syllable truncation was followed by either a song stop (light gray) or a motif restart (dark gray). In only a small percentage of cases (2.8%, all birds and all stimulus intensities) was syllable truncation not followed by any song-level effect (white).
When viewed as a percentage of affected songs at each stimulus intensity, a significant number of syllable-level effects were seen in all birds. Even at the lowest intensity (15 μA), stimulation caused a significant number of syllable truncations or distortions when compared with unstimulated control songs (p < 0.001, G test) (Fig. 3B). Stronger stimulation intensities resulted in a larger proportion of syllable-level effects. For instance, 45 μA stimulation caused noticeable spectral or amplitude distortion in 42.3% of songs (289 of 684 songs; p < 0.001, G test) and truncation in another 25.0% (171 of 684 songs; p < 0.001, G test) (Table 1).
Effect of 45 μ A stimulation on song output by area
The second category of effects that were analyzed consisted of song-level effects (Fig. 2B), which represents the percentage of songs in which stimulation halted (song stop) or restarted (motif restart) the entire motif. Song-level effects (restarting and stopping) were seen in more than half of the song set (65.2%; n = 1783 of 2735 songs in 4 birds) (Fig. 3A). Similar to the analysis of syllable-level effects, we also calculated the percentage of all songs for all birds that showed song-level effects at each stimulus intensity, ranging from 15 to 60 μA. These percentages were compared with a control set of songs (Fig. 3C). A significant number of song-level effects were seen at all stimulus intensities. For example, 45 μA stimulation caused restarting in 40.8% of songs (279 of 684 songs in 4 birds; p < 0.001, G test) and stopping in 33.6% (230 of 684 songs in 4 birds; p < 0.001, G test).
Syllable-level and song-level effects often occurred together in the same song after stimulation. When syllable truncation occurred, it almost always preceded a song effect (Fig. 3D). Of all truncations produced at 45 μA, 63.2% of these preceded a restart, and 33.3% preceded a song stop (n = 108 and 57 instances, respectively, of 171 truncates in 4 birds at 45 μA). Only 3.5% of truncates at this level occurred in the absence of any song effect. Similar distributions were seen at all stimulus levels and all birds (Fig. 3D). Song-level effects followed syllable distortion less often than truncation. Of all the songs that showed syllable distortion at 45 μA (n = 289 song in 4 birds), 33.9% (n = 98) preceded a restart, and 41.5% (n = 120) preceded a complete song stop. Similar values were seen at all stimulus levels. Across all intensities, restarts were more commonly observed after syllable truncation (63.4%; n = 313 of 494 songs) than they were after syllable distortion (38.0%; n = 348 of 915 songs). Conversely, song stops were more commonly observed after distortion (36.6%, n = 335 of 915 songs) than after syllable truncation (33.8%; n = 167 of 494 songs). These distributions of song-level effects after distortion and truncation were significantly different from one another (p < 0.001, χ2 test).
Syllable truncation occurs with a fixed latency relative to stimulation
Latency to syllable truncation was measured as an indicator of how rapidly stimulation could affect song production. Stimulation in HVC caused truncation to occur with a latency of 74.4 ± 15.6 ms (n = 111 songs in 4 birds) (Fig. 4C). Truncations occurred at any time throughout syllables, without respect to syllable or note boundaries, as revealed by comparing the lengths of stimulated syllables to the distribution of normal unstimulated syllable lengths (Fig. 4). As shown in Figure 4A for one sample syllable, the length of control syllables was tightly clustered around the mean syllable length. In contrast, the length of stimulated syllables was broadly distributed over a wide range with no obvious clustering at either note transitions or at the end of the syllable. In Figure 4B, syllable truncation times are compiled from each of four birds (one syllable per bird) with syllable lengths and stimulation times normalized to the mean length of each bird's respective syllable. Similar to the distribution shown in Figure 4 A, truncation times for this combined dataset are distributed throughout the stimulated syllables. Again, truncations occurred with a fixed latency after stimulation, and the distribution of syllable truncation lengths was correlated with the distribution of stimulation times. For the syllables analyzed in Figure 4 B, stimulation times and the times of truncation were correlated (r = 0.58), which was significant when compared with a time-shuffled control (z = 2.63; p < 0.005), and was time shifted by 80 ms (using 10 ms bins). This time shift was similar to the mean latency to truncation for all syllables in all birds (74.4 ± 15.6 ms; n = 111 songs in 4 birds) (Fig. 4C). As expected, the lengths of unstimulated control syllables were tightly clustered around the mean lengths of the syllables. These observations indicate that HVC stimulation can exert an effect at any point during song production, and that these effects occur with a predictable latency that is unrelated to expected syllable boundaries.
To investigate whether truncation latency varied as a function of stimulation delivery time within a syllable, we correlated stimulation time with truncation latency (Fig. 4D). We found that microstimulation produced truncations with a similar delay regardless of when it occurred in the syllable (linear regression, R2 = 0.067). Even stimuli that were delivered in the silent interval that preceded a syllable caused the syllable to truncate with a latency that was similar to that observed when stimuli were delivered during active vocalization.
Finally, to examine whether stimulation suppressed the ongoing syllable but left the overall timing of the next syllable intact, we examined the relationship between the duration of the truncated syllable and the length of the silent interval between the offset of that syllable and the onset of the following syllable. Because the subset of truncations analyzed here were only those that preceded a motif restart, this next syllable was usually an introductory note or the first syllable of a new motif. If the timing of the restart were fixed and independent of stimulation and truncation time, silent interval length should increase when the syllable is shortened by stimulation, creating an inverse linear correlation between the truncated syllable length and the silent interval length. However, we found no significant correlation between these values in any of the four birds tested (r = 0.15 ± 0.21, Pearson's r test; n = 109 songs in 4 birds). Duration of the silent interval tended to be variable and had a mean latency of 120.6 ± 46.6 ms (n = 109 songs in 4 birds). These data suggest that HVC stimulation truncates syllables with a fixed latency but that the latency to the onset of the next syllable is not fixed during restarts of the motif.
In a very small number of cases (n = 6 of 2191 songs in 4 birds; 30–60 μA stimulation), truncation was not followed by a song-level effect but rather by continuation of the ongoing motif with the production of the next syllable in the motif sequence. In these instances, the length of the truncated syllable was inversely correlated with the silent interval length (r = –0.59, Pearson's r test; n = 6 songs in 4 birds), indicating that, when syllables were truncated to shorter lengths, the delay to the next syllable was correspondingly longer. Consistent with this relationship, the time from the start of truncated syllables to the start of following syllables was not significantly different from the interval measured in unstimulated matched syllable pairs (p = 0.39, paired Student's t test). These results suggest that the timing from one syllable to the next syllable in the motif sequence is preserved in the rare cases when stimulation-induced syllable truncation does not precede song-level effects.
Stimulation of HVC alters respiratory pressure during song production
The structure of each syllable of a bird's song is associated with a stereotyped respiratory pattern that can be measured by examining air sac pressure (Franz and Goller, 2002). In zebra finches, phonation is produced almost exclusively during expiration (with rare exceptions), and inspiration is predominantly associated with silent intersyllable intervals (Suthers et al., 1999). To assess the latency with which stimulation affects the underlying respiratory pattern, we performed unilateral HVC stimulation during normal respiration and singing and measured air sac pressure as an indicator of activity changes in the brainstem respiratory network.
In a first set of experiments, we asked whether HVC activity could directly influence the respiratory network, as has been suggested previously (Manogue and Paton, 1982). Electrical stimulation (six biphasic pulses, 400 μs long) was delivered to HVC in unrestrained awake birds every 10 s during quiet respiration, and air sac pressure was recorded with an implanted cannula attached to a pressure transducer (see Materials and Methods). Multiple air sac pressure traces were averaged and aligned with the start of stimulation. Unilateral HVC stimulation caused a brief increase in air sac pressure, regardless of whether stimulation occurred during the inspiratory or the expiratory phase. Stimulation-evoked increases in air sac pressure were observed in all birds tested, and data for one bird is shown in Figure 5A. Stimulation at 45 μA resulted in a positive deviation in the pressure trace that was 33.4% of the maximum normal pressure range (measured from maximum during expiration to minimum during inspiration; n = 66 stimulations). Stimulation-evoked changes in the other two birds were 41.2 and 114.3%, respectively.
Despite differences in intensity, HVC stimulation produced air sac pressure changes with a similar time course in all three birds. Mean time to reach the peak of the pressure increase after stimulation was 30.4 ± 2.7 ms, while the time to half the peak was 22.3 ± 1.5 ms (n = 94 stimulations in 3 birds). To better estimate the earliest deviation from baseline pressure levels, we measured the duration between stimulus onset and the time it took the pressure trace to deviate from the mean of the baseline trace by 2 SDs. Measured this way, significant changes in the pressure pattern were observed as soon as 17.6 ± 2.2 ms after stimulation onset (n = 94 stimulations in 3 birds; for details, see Materials and Methods).
During singing, HVC stimulation resulted in rapid changes in respiratory pressure (Fig. 5B). In cases when stimulation caused the ongoing syllable to truncate, there was an accompanying rapid (within 20–30 ms) initial decrease in pressure that was followed by a return to baseline pressure levels (within 60–80 ms). In some cases, a similar initial decrease in pressure was seen without truncation or song-level effects, resulting only in syllable distortion. The temporary increases in pressure described above for quiet respiration could not be detected after stimulation during song. Such small increases in pressure might have been obscured in the background of the 5- to 10-fold increase in pressure levels normally seen during singing.

Characteristics of syllable truncation after stimulation. A, Relationship of stimulation times to syllable truncation in a sample syllable. In the top panel, each tick mark represents the time of stimulus delivery to an instance of the syllable. The distribution of syllable lengths resulting from these stimulations (middle panel) illustrates that the length of truncated syllables is evenly distributed (gray bars) throughout the duration (∼170 ms) of a normal unstimulated syllable. The y-axis represents the percentage of all truncated syllables analyzed for this bird. For comparison, the distribution of syllable lengths for unstimulated instances of the syllable is shown below (black), along with typical times for the note transition (white) in that syllable. The mean time from stimulus onset to truncation (truncation latency) is shown as a dotted line for reference and represents the expected delay from stimulation time to truncation time. The bottom two traces represent, respectively, the spectrogram and acoustic waveform of the syllable. B, Compiled distribution of truncation times for all birds. For each bird, one long syllable (ranging from 102 to 207 ms) was chosen to analyze the distribution of truncated syllable lengths. As described above, stimulation times are shown in the top panel as thin lines and are normalized such that 100% represents the mean length of each syllable. The length of stimulated (gray) and unstimulated (black) syllables is plotted in the bottom panel. C, Histogram of syllable truncation latencies after stimulation in HVC, for a total of 111 syllables representing a sample syllable for each of four birds. Bin size, 10 ms. D, Truncation latency as a function of time of stimulation. Truncation latencies are shown for four birds, plotted against the time of stimulation shown as the percentage of syllable length. As the trend line indicates, there is no correlation between the time of stimulation and truncation latency.
To estimate the latency with which the respiratory pattern changed, we examined syllable truncations and distortions in detail and compared respiratory traces during stimulated syllables to an average profile of respiratory activity for the same unstimulated syllable (for details, see Materials and Methods). An example is shown in Figure 5B, in which a respiratory pressure trace during stimulation (black line) is shown overlaid on the mean trace for this syllable (gray line). The light gray area around the mean trace represents a distance of 2 SDs from the mean. Divergence of the stimulation trace by more than 2 SDs from the mean provides a conservative estimate of the latency of stimulation effect during singing. Using this criterion, the mean latency to cause a deviation in air sac pressure was 27.3 ± 7.2 ms (n = 35 songs in 3 birds). This latency is ∼10 ms longer than the time it takes to alter air sac pressure during normal respiration.

Effects of HVC stimulation on air sac pressure. A, During quiet respiration, HVC stimulation caused a small, brief increase in air sac pressure. Each trace represents the average of multiple (55–96) air sac pressure traces, aligned with the time of unilateral HVC stimulation at four different current intensities (15, 30, 45, and 60μA). All traces represent stimulation in one bird. The white box starting at 0 ms represents the period of stimulation. B, During song, stimulation causes a short-latency deviation from the stereotyped respiratory pattern. The dark gray trace is the mean of multiple (n = 18) air sac pressure traces recorded during production of the three sequential syllables in the bird's motif. The light gray zone surrounding this unstimulated air sac pressure trace represents 2 SDs from the mean trace. The black line in this figure represents the air sac pressure trace recorded for a stimulated syllable. Stimulus time is represented by the double dotted lines. Sample spectrograms for both the unstimulated and stimulated syllables are shown at the bottom. In this example, stimulation resulted in a syllable truncation. C, Example of a stimulation-induced change in air sac pressure that does not result in a song-level effect. The top two traces represent, respectively, the acoustic waveform and the air sac pressure trace of two sequential motifs. A stimulus was delivered during the production of the syllable B. Higher magnification of this syllable is shown in the bottom panel, with the stimulus shown by the arrow and the white box. The envelope of the air sac pressure trace is similar to that described above, with the stimulated syllable in black and the control trace in gray. The gray zone surrounding the control trace represents 2SDs from the mean. In this example, the song shows no song-level effects despite a clear decrease in air sac pressure in the stimulated syllable. D, Relationship between stimulus-induced changes in air sac pressure and song-level effects. The presence or absence of song-level effects was measured in the subset of cases in which stimulation caused a significant change in air sac pressure. The majority of syllables that showed a significant change in air sac pressure after stimulation were also followed by either motif restarts or song stops. However, some syllables (10 of 46 syllables) failed to show any song-level effects despite the stimulus-induced decrease in air sac pressure.
These sorts of changes in air sac pressure outside the SD line were frequently associated with the eventual truncation of the syllable, and this was often followed by stopping or restarting of the song. However, we observed a number of cases in which there was a change in pressure that was not accompanied by a syllable truncation or by song-level effects (Fig. 5C). Although such examples only occurred a small proportion of the time (21.7% of songs with significant air sac pressure deviation; n = 10 of 46 songs in 3 birds) (Fig. 5D), these results argue that intermediate changes in air sac pressure can occur without resulting changes to the song pattern.
Temporal sequence of events after HVC stimulation
To investigate the sequential order by which HVC stimulation causes changes in the song pattern, we compared the latency to acoustic truncation with the latency to air sac pressure changes, which provides an indirect representation of the effect on the respiratory network. As expected, significant deviation from normal air sac pressure after HVC stimulation (27.3 ± 7.2 ms; n = 35 songs in 3 birds) was significantly shorter than the latencies for complete acoustic truncation (74.4 ± 15.6 ms; n = 111 songs; F = 61.3; df = 1, 5; p < 0.01, NANOVA) (Fig. 6). These results imply that, on average, suppression of the respiratory network precedes changes in the acoustically observed truncation of the syllable.
It has been shown previously that truncation of syllables and the corresponding song-level stops or restarts resulting from HVC stimulation are accompanied by a short-latency suppression of neural activity within the HVC contralateral to the stimulation site (Vu et al., 1998). We compared latencies to air sac pressure change (which usually accompanied song-level effects) with the time it takes to suppress contralateral HVC neural activity during a stop or restart of the song. The dataset used for this comparison is the same as presented in a previous study (Vu et al., 1998). As shown in Figure 6, the latency to changes in air sac pressure (27.3 ± 7.2 ms; n = 35 songs in 3 birds) was significantly shorter than the latency for contralateral HVC reset (36.1 ± 12.8; n = 20 songs; p < 0.01, Student's t test) (Vu et al., 1998). This observation suggests that, on average, HVC stimulation affects the brainstem respiratory centers before it affects the contralateral HVC.
Stimulation within RA interrupts song
Because stimulation causes a short-latency perturbation of respiration, we asked whether stimulation within the dorsal region of RA (dRA) [which projects preferentially to respiratory-vocal nuclei in the brainstem (Wild, 1993a, 1997)] could disrupt song patterns in ways similar to those produced by stimulation in HVC. We implanted stimulating electrodes within the dorsal-most 100–200 μm of RA in normal adult birds and delivered brief microstimulation pulses to this area during song production. For comparison, electrodes were also implanted in the ventral region of RA (vRA) of these same birds. The ventral portion of RA projects predominantly to nXIIts (Vicario, 1991).
We found that low-level microstimulation in the dorsal RA altered song in ways strikingly similar to HVC stimulation, in terms of both syllable-level effects (Fig. 7A, left panel) and song-level effects (Fig. 7B, left panel). Previous studies (Vu et al., 1994) failed to elicit song-level effects in RA with less than seven 400 μs pulses at 45 μA. We nonetheless found that, in dorsal RA, stimulation with as few as five pulses at 30 μA produced syllable-level effects (truncation or distortion) 53.0% of the time (n = 349 of 659 songs in 4 birds; p < 0.001 compared with control songs, G test) and song-level effects (stop or restart) 50.5% of the time (n = 333 of 659 songs in 4 birds; p < 0.001, G test). Surprisingly, the same stimulation parameters in ventral RA of these same birds also produced syllable-level effects 41.2% of the time (n = 145 of 352 songs in 4 birds; 30 μA intensity; p < 0.001, G test) and song-level effects 47.7% of the time (n = 168 of 352 songs in 4 birds; p < 0.001, G test). Stimulation at 45 μA produced similar results for syllable-level (p < 0.001, G test) (Fig. 7A, right panel) and song-level (p < 0.001, G test) (Fig. 7B, right panel) effects in both vRA and dRA. Compared with control songs, a significant number of syllable-level and song-level effects were seen in both the ventral and dorsal RA for all stimulus levels tested (p < 0.001, G test) (Fig. 7, Table 1). A significantly greater number of syllable-level effects were seen for the dorsal stimulation group compared with the ventral group (n = 593 songs in 3 birds; p < 0.001, G test). No significant difference was seen for song-level effects (n = 593 songs in 3 birds; NS, G test) between dRA and vRA. Although the stimulation techniques used in this study were nearly identical to those of Vu et al. (1998), the electrodes used by Vu et al. (1994) were of lower impedance and different construction (Vu et al., 1994) and may account for some of the differences seen here.

Latency to truncation after HVC stimulation at different levels of the song motor control system. Latency to syllable truncation (Acoustic truncate) was significantly longer than the latency to air sac pressure change (Air sac pressure change). This air sac pressure change latency is also shorter than the latency to suppression of premotor activity in the contralateral HVC (Contra HVC neural reset). Data for this last category were the same as those reported in a previous study (Vu et al., 1998).

Effects of unilateral stimulation within RA during song. A, Percentage of songs with syllable-level effects resulting from RA stimulation. On the left, the percentage of syllable-level effects combined from both dorsal and ventral RA stimulation is shown for four different current intensities. On the right, syllable-level effects at 45 μA are compared between the dorsal and ventral regions of RA. B, Percentage of song-level effects resulting from RA stimulation. On the left, effects from ventral and dorsal stimulation are combined. On the right, song-level effects are shown separately for 45μA stimulation in the dorsal and ventral regions of RA. All categories are compared with percentages collected from a control set of songs (CONT) with no stimulation (*p < 0.001, G test).
The similarity of effects observed in dRA and vRA might suggest that both subdivisions of RA contribute in similar ways to song motor pattern generation. Alternatively, some of the present results (particularly those for higher stimulation levels) might be the result of antidromic activation of HVC neurons, some of which are known to project to both dRA and vRA (Kittelberger and Mooney, 1997). It may also be possible that stimulation within dRA might antidromically or orthodromically affect the ventral portion of the nucleus, a likely possibility in view of the extensive interactions within different parts of RA (Spiro et al., 1999). These results alone can therefore not conclusively address the differential contribution of the ventral and dorsal portions of RA to motor pattern generation. To examine this issue more directly, we performed microstimulation within two brainstem nuclei that receive projections from RA. One of the targeted regions was a nucleus within the respiratory network, specifically the inspiratory-related nucleus PAm, which receives projections from the dorsal region of RA and which, in addition to its spinal projections to motoneurons innervating inspiratory muscles, projects bilaterally back into the forebrain song system via the thalamic nucleus Uva (Wild, 1993a,b, 1997, 2004b; Reinke and Wild, 1998). The second region was nXIIts, which receives projections from the ventral region of RA and controls the muscles of the syrinx (Vicario, 1991). Unlike PAm, nXIIts possesses no known projections back into the forebrain song system.
Stimulation in nXIIts affects acoustic structure but not song pattern
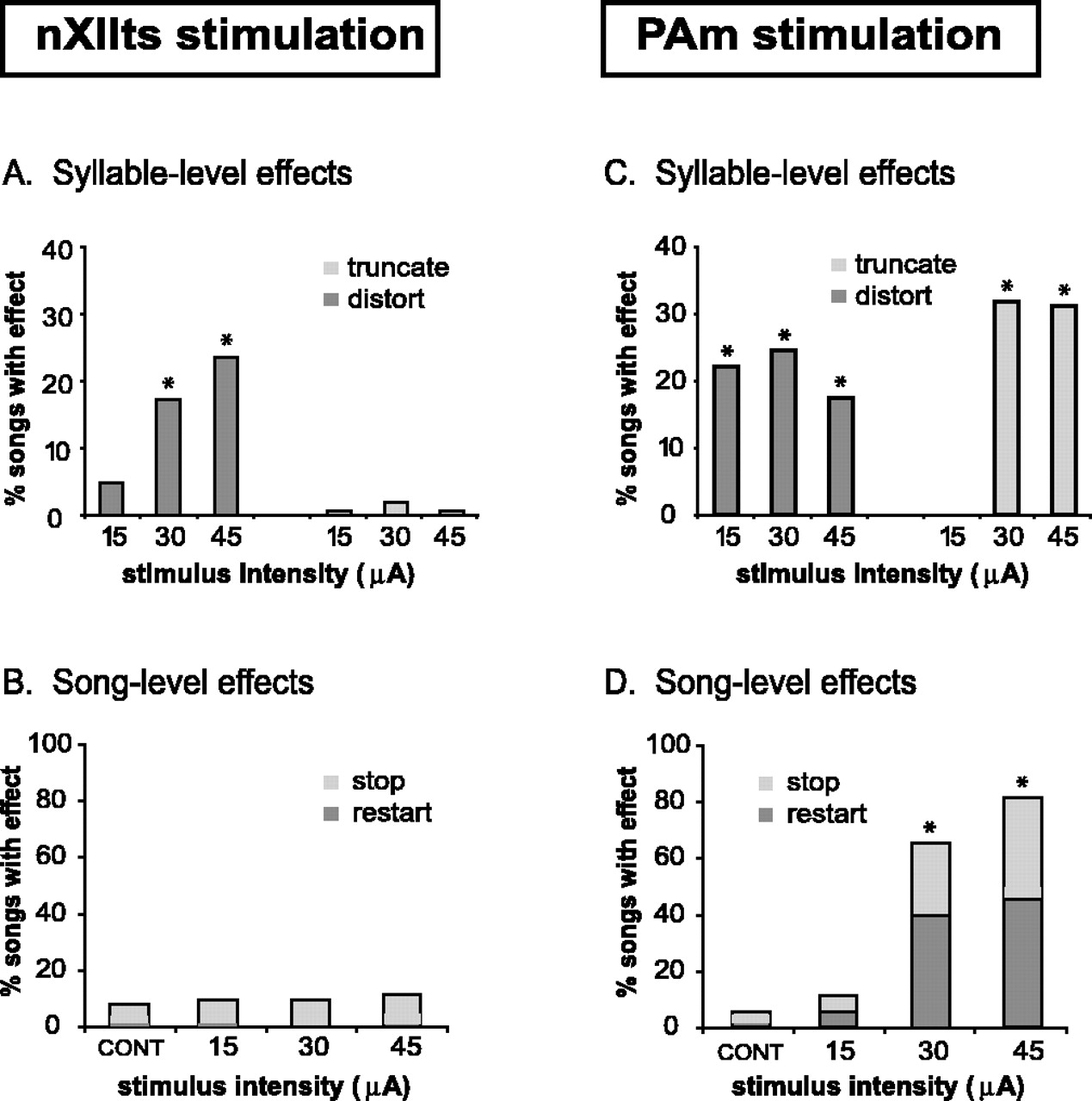
Because nXIIts innervates the syrinx, which controls the acoustic structure of vocalizations, we expected to see distortions of syllables but not truncations or song-level effects (stop or restart of the song) from nXIIts stimulation. Consistent with this hypothesis, we found that stimulation within nXIIts did not cause a significant number of syllable truncations at 15, 30, and 45 μA (0.5% for 15 μA, 1.9% for 30 μA, and 0.5% for 45 μA; n = 607 songs in 3 birds; NS, G test). Stimulation at 15 μA also did not cause a significant number of syllable distortions (4.7%; NS, G test), but, as expected, stimulation at 30 and 45 μA did (17.0 and 23.2%, respectively; p < 0.001, G test) (Fig. 8A, Table 1). Stimulation at 15, 30, or 45 μA did not cause any significant song-level effects (NS, G test) (Fig. 8B). For example, 45 μA stimulation only preceded restarts 1.5% of the time (n = 3 of 203 songs) and stops 9.9% of the time (n = 20 of 203 songs) compared with 0.6 and 7.3%, respectively, in the 0 μA control set (n = 164 songs in 3 birds; NS, G test). Both syllable-level and song-level effects were nevertheless observed when stimulation as high as 60 μA was used. Distortion followed 60 μA stimulation 36.0% of the time (82 of 228 songs), although truncation followed only 0.9% of the time (2 of 228 songs). Restarts followed 60 μA stimulation 2.2% of the time (5 of 228 songs), and stops followed 60 μA stimulation 11.8% of the time (27 of 228 songs, song-level effects; p < 0.01, G test). These effects were not unexpected given the possibility of antidromic activation of the respiratory network through some of the projections known to exist between brainstem respiratory nuclei and nXIIts (Reinke and Wild, 1998; Sturdy et al., 2003). Overall, these observations that brief stimulation in nXIIts causes syllable distortions without any obvious effect on syllable duration or song pattern are consistent with a role for this nucleus in determining acoustic structure but not overall temporal pattern. Thus, the network of nuclei controlling motor program generation for song is unlikely to include the output pathway leading to the vocal apparatus.

Effects of unilateral stimulation within nXIIts (left panels) and PAm (right panels) during song. nXIIts stimulation (A, B). A, Syllable-level effects. Stimulation in this structure caused significant distortion of the ongoing syllable (dark gray) without any significant syllable truncation (light gray). B, Song-level effects. Stimulation in nXIIts did not cause any significant amount of song stops (light gray) or motif restarts (dark gray). PAm stimulation (C, D). C, Syllable-level effects. Stimulation in PAm caused both truncation of the ongoing syllables (light gray) and acoustic or spectral distortion of the stimulated syllable (dark gray). D, Song-level effects. In contrast to nXIIts, stimulation in PAm caused a significant number of song stops (light gray) and motif restarts at 30 and 45 μA (dark gray). Syllable-level effects are shown for three different current levels (A, C) and are compared with a control set (data not shown). In the song-level graphs (B, D), the control set of songs with no stimulation is labeled CONT (*p<0.001, G test).
Stimulation within the respiratory network interrupts song
In contrast to nXIIts stimulation, we found that stimulation within PAm resulted in stopping and restarting of song at rates comparable with those seen in HVC and RA. A significant number of syllable-level effects were also seen for all stimulation intensities (Fig. 8C), although, at 15 μA, this was attributable to distortions only. At 30 μA, stimulation in PAm caused syllable distortion 24.4% of the time and truncation 33.3% of the time (n = 19 and 26 songs, respectively, of 78 songs in 3 birds; p < 0.001, G test). At 45 μA, the syllable-level effects were similar (18.8% distorts, 32.8% truncates; n = 12 and 21 songs, respectively, of 64 songs in 3 birds; p < 0.001, G test). At the song level (Fig. 8D), 30 μA stimulation in PAm caused restarts 39.7% and stops 25.6% of the time (n = 31 and 20 songs, respectively, of 78 songs in 3 birds; p < 0.001, G test). Stimulation of 45 μA produced a slightly larger number of song-level effects (45.3% restarts, 35.9% stops; n = 29 and 23 songs, respectively, of 64 songs in 3 birds; p < 0.001, G test). The song-level effects seen in PAm are consistent with a role for this nucleus in contributing to the structure of motifs or at least providing important input back into the forebrain motor network.
To measure the temporal relationship between effects caused by HVC and PAm stimulation, we examined latencies to acoustic syllable truncation after stimulation in both structures. Latency to truncation was significantly shorter for PAm stimulation than for HVC. The mean latency was 51.2 ± 6.9 ms for PAm (n = 27 songs in 3 birds) compared with 74.4 ± 15.6 ms for HVC stimulation in normal birds (n = 111 songs in 4 birds; F = 171.2; df = 1, 5; p < 0.01, NANOVA). A summary of stimulation effects on song and associated latencies to truncations for different nuclei in the song motor pathway is shown in Table 1.
Stimulation within the respiratory network causes temporary vocal suppression
Stimulation in the inspiratory-related nucleus PAm at 30, 45, or 60 μA often caused a temporary suppression of vocal output that was distinct from syllable truncations. Such temporary suppression was observed in 19.2% of songs (54 of 281 songs in 3 birds) and was characterized by a drop in acoustic amplitude to near baseline levels. This was followed by a return to normal singing volume before the end of the stimulated syllable (Fig. 9). This form of distortion was rarely seen with stimulation in other areas and was possibly attributable to the direct control by PAm of inspiratory muscle activity. On average, acoustic amplitude suppression occurred 22.3 ± 6.6 ms after stimulation onset and lasted 12.7 ± 7.0 ms. In some cases (27.8%, 15 of 54 songs with suppression), this temporary event was followed by a second and final return to baseline, resulting in a truncated syllable. Although temporary suppression was often followed by song-level effects (18.5% restarts and 31.5% stops; 10 and 17, respectively, of 54 songs with suppression) (Fig. 9B), we observed many cases in which stimulation caused temporary acoustic suppression without any song-level effect (50.0%; 27 of 54 songs) (Fig. 9C). Conversely, in a majority of the cases in which song-level effects occurred, no temporary acoustic suppression was seen (79.1%; 102 of 129 songs with song-level effects). These results indicate that the suppression and song-level effects can occur independently and that neither is a prerequisite for the other.
Discussion
We used brief microstimulation to perturb ongoing premotor activity in discrete nuclei along the song motor pathway. Stimulation caused two categories of perturbation: (1) syllable-level effects (syllable distortion and truncation), and (2) song-level effects (song stops and motif restarts). Song-level effects represent an interruption of the song motor program and thus reflect a role for the stimulated target nucleus in the generation of the song temporal structure. We show that perturbation of premotor activity in nuclei within the recurrent song system motor network, but not in output structures such as nXIIts, will disrupt the ongoing song motor sequence. These results support a model whereby the brainstem vocal-respiratory nuclei form an integral part of the song motor programming network by providing timing signals and information about the current respiratory state to song control nuclei in the forebrain.
Mechanism of HVC stimulation-evoked song interruption
Stimulation in HVC during song frequently caused complete truncation of ongoing syllables, which was usually followed by disruption in the ongoing song sequence. The latency to acoustic truncation of syllables was longer and more variable (74.4 ± 15.6 ms) than the latency to air sac pressure disruption (27.3 ± 7.2 ms). This suggests that stimulation in HVC disrupts the vocal respiratory system, which then causes a decrease in air sac pressure. This suppression of respiratory drive then ultimately leads to truncation of the ongoing syllable. The difference in latency between acoustic truncation and air pressure change is likely caused by continued premotor activity in the unperturbed contralateral HVC. Previous studies (Vu et al., 1998) have shown that premotor activity in the contralateral HVC remains active for 30–50 ms after stimulation before it becomes suppressed to baseline levels. During this period, contralateral HVC output is thus able to effect sufficient syringeal muscle activity to produce structured acoustic output during the remaining deflation of residual pressure in the air sacs. These results agree with our observations that latency to acoustic truncation after HVC stimulation is much shorter in birds forced to sing with only one hemisphere than it is in intact birds (Ashmore et al., 2004). Together, the timing of these different perturbations after stimulation in HVC are consistent with a model in which brief interruptions of the ongoing premotor activity in HVC will cause disruption, and ultimately suppression, of the ongoing respiratory pattern, which then causes a perturbation of the song motor program.
Song representation in HVC
During a normal song bout, zebra finches sing multiple motifs and end most of their songs on the same syllable each time. In a few cases, however, birds will abort song prematurely, ending on uncharacteristic syllables. When this occurs, either spontaneously or attributable to manipulations such as strobe lights (Cynx, 1990) or delayed auditory feedback (Cynx and Von Rad, 2001) (J. Cynx, personal communication), motifs are usually disrupted at syllable boundaries, in which respiration switches from expiration to inspiration. Based on these observations, syllables and their corresponding respiratory cycle have been suggested to make up the basic “motor unit” of song production (Suthers and Margoliash, 2002), and previous studies have suggested that HVC might encode song at the syllable level (Yu and Margoliash, 1996).
One possible interpretation of such a motor unit representation at the level of HVC is that ongoing network activity within this nucleus generates “punctuated” instructions regarding syllable identity and transmits these to downstream nuclei at the beginning of each syllable. Stimulation in HVC should therefore cause songs to truncate at syllable boundaries regardless of when the stimulation is delivered relative to these boundaries. Our results are inconsistent with this scenario because HVC stimulation causes syllables to truncate after a fixed delay regardless of when in the syllable the stimulus is delivered. Rather than coding syllables as a motor unit, sending instructions to RA at syllable boundaries, HVC appears to code syllables at a finer temporal scale. These findings provide functional support for the idea that neural output from HVC to RA encodes song on a moment-to-moment level, with small populations of projection neurons each representing a single window of time in the song (Hahnloser et al., 2002). Although syllables may still be represented as a basic motor unit of song production at some level in the song motor pathway, our results suggest that this representation may not exist in HVC.

Changes in acoustic amplitude pattern are not sufficient to cause song-level effects. A, Amplitude envelope of an unstimulated control syllable. The top panel represents a motif from an unstimulated control song shown as an amplitude waveform. In the bottom half of the panel, a representative syllable (syllable C, solid line) has been rectified and smoothed. It is overlaid over the mean envelope of that same syllable (dotted line) to show the stereotyped nature of the acoustic envelope of unstimulated syllables. B, Biphasic change in acoustic amplitude pattern after PAm stimulation leading to syllable truncation. Stimulation (indicated by vertical arrow) in PAm often caused a rapid and temporary decrease (highlighted in gray) in the acoustic amplitude of the stimulated syllable (solid line). As shown in this example, this was sometimes followed by a temporary increase in amplitude of the syllable acoustic envelope. The temporary suppression preceded syllable truncation, which is exemplified by a second and final decrease of acoustic amplitude to baseline levels. Syllable truncation was then followed by a song-level effect (restart). This biphasic change in acoustic pattern after PAm stimulation was not observed in the other structures that were stimulated. The average envelope of the unstimulated control syllable C is shown as a dotted line. C, Biphasic change in acoustic amplitude pattern that does not lead to song-level effects. In this example, stimulation caused a temporary suppression (highlighted in gray) of the acoustic envelope of syllable C without any ensuing song-level effect. All examples in A–C were for the same syllable in different songs in the same bird. Current intensity of PAm stimulation was 45μA in both B and C. D, Relationship between acoustic amplitude changes and song-level effects. In half (27 of 54) of the instances of temporary suppression, song-level effects (song stop and motif restart) followed. In the rest (27 of 54), no song-level effects were seen.
Interruption of the song motor pattern by stimulation in RA and the brainstem respiratory network
In contrast to a previous study (Vu et al., 1994), we found that stimulation in RA was able to truncate syllables and alter the song pattern at low current intensities. This was true for stimulation in the dRA, which projects primarily to respiratory nuclei and to the DM (vocal dorsomedial nucleus of the intercollicular complex), as well as the vRA, which projects to the hypoglossal nucleus (nXIIts), which in turn innervates the syrinx (Fig. 1). The reasons for the difference between our study and that of Vu et al. (1994) are not clear. It seems unlikely that the lowest stimulation intensities would exert their effects via antidromic activation of HVC. Nonetheless, we addressed this possibility, as well as the potential confound created by intrinsic connections within RA (Spiro et al., 1999), by implanting stimulating electrodes in two of the known targets of RA, nXIIts, and the inspiratory premotor nucleus PAm.
Stimulation in nXIIts was not effective in altering the song temporal structure, suggesting that the pathway from vRA to nXIIts to the syrinx is not part of the motor pattern-generating network. In contrast, stimulation in PAm caused syllable and song-level effects that were similar to those observed in HVC and RA. Given that nuclei in the vocal-respiratory network, in particular PAm and DM, send bilateral recurrent projections back into the forebrain, we propose that these nuclei contribute to song motor pattern generation through activity along these projections.
In addition to causing syllable truncation, stimulation in PAm also caused a short-latency (22.3 ms) and brief reduction in amplitude of the vocal output in a subset of songs. This temporary disruption of amplitude was unlikely to be related to the song-level effects seen from PAm stimulation, because the majority of song stops and restarts were observed in the absence of any change in acoustic amplitude. Conversely, in half of the cases in which temporary suppression did occur, no song-level effects were observed. Change in respiratory drive is therefore neither necessary nor sufficient to produce song-level effects. The presence of two independent effects is consistent with the observation of two distinct populations of neurons in PAm, a bulbospinal premotor group mediating inspiratory control, and another group whose axons ascend and terminate bilaterally within Uva (Reinke and Wild, 1998; Wild, 2004b). The existence of two separate events after PAm stimulation suggests that song-level effects are not simply a product of respiratory feedback resulting from sudden changes in air sac pressure or volume. This is consistent with previous work showing that artificially induced changes in air sac pressure, by injections of air into the cranial thoracic air sac, cause a rapid readjustment of the pressure pattern without any effect on syllable order or motif structure (Suthers et al., 2002). It is important to note that these observations do not by themselves rule out a possible role for somatosensory feedback in the process of song generation or the process of interruption caused by stimulation.
Recurrent loop models for song production
Many models of complex or learned motor production are framed in a hierarchical and linear manner, with the forebrain guiding the activity of brainstem nuclei, which in turn act primarily as output channels (Jürgens, 2002; Krauzlis, 2004). In the avian song system, for example, forebrain song control nuclei HVC and RA are thought to generate the motor commands for the different brainstem nuclei that, respectively, control respiration and the syrinx (Suthers and Margoliash, 2002; Fee et al., 2004). Despite its intuitive appeal, a hierarchical view of motor control in the avian song system fails to explain our current results as well as the observed synchronization of premotor activity between left and right HVC (Schmidt, 2003). Given the functional and anatomical organization of the song system, song motor control can be alternatively explained with a recurrent flow of information through the song system. Although HVC is likely to contribute to song pattern generation (Solis and Perkel, 2005), each HVC in this alternative model is ultimately dependent on the activity in the vocal-respiratory network [nucleus retroambigualis (RAm), PAm, and DM], which would in turn be dependent on inputs from the forebrain song control nuclei RA and HVC.
Although the nature of song-related activity in the vocal-respiratory network is presently unknown, sensory information derived from both the lung-air sac system and the syrinx may form functionally important inputs to this network. Pulmonary and syringeal afferents terminate separately in adjacent regions of the parasolitary subnucleus of the nucleus of the solitary tract, which, in turn, projects on PAm (Wild, 2004b). Inputs from the vocal-respiratory network to the forebrain could serve the role of a clock (Williams, 1989; Williams and Vicario, 1993), driving or modulating HVC activity at syllable or subsyllable intervals. This clock would serve to inform forebrain song control nuclei of the current respiratory state, to maintain a tempo that is consistent with the physical constraints of the respiratory system, and to synchronize the two hemispheres throughout song production. Although brainstem vocal-respiratory structures undoubtedly serve as output pathways for the song system, our results suggest that the recurrent circuit provides critical timing information to song control nuclei in the forebrain and therefore forms an integral part of the song pattern-generating network.
Footnotes
This research was supported by National Institutes of Health Grant RO1 DC006102 and the Alfred P. Sloan Foundation. We thank Mark Bourjaily for assistance with song analysis and scoring, Josh Gold and Paul Nealen for useful comments and help with statistics, Brent Cooper for technical assistance with air sac pressure recording, Peter Petraitis for statistical consultation, Ofer Tchernichovski for technical assistance with the Sound Recorder and Sound Analysis software, John Andrews-Labenski for technical assistance with microstimulation, and Fabiana Kubke for detailed constructive commentary on this manuscript.
Correspondence should be addressed to Marc F. Schmidt, 312 Leidy Laboratories, Department of Biology, University of Pennsylvania, Philadelphia, PA 19104-6018. E-mail: marcschm{at}sas.upenn.edu.
DOI:10.1523/JNEUROSCI.1668-05.2005
Copyright © 2005 Society for Neuroscience 0270-6474/05/258543-12$15.00/0





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}