

Abstract
During development, a specific subset of ventral spinal cord precursors called pMN cells produces first motor neurons and then oligodendrocyte progenitor cells (OPCs), which migrate, divide and differentiate as myelinating oligodendrocytes. pMN cells express the Olig2 transcription factor and Olig2 function is necessary for formation of spinal motor neurons and OPCs. In the hindbrain and midbrain, distinct classes of visceral, branchiomotor and somatic motor neurons are organized as discrete nuclei, and OPCs are broadly distributed. Mouse embryos deficient for Olig2 function lack somatic motor neurons and OPCs, but it is not clear whether this reflects a common origin for these cells, similar to spinal cord, or independent requirements for Olig2 function in somatic motor neuron and OPC development. We investigated cranial motor neuron and OPC development in zebrafish and found, using a combination of transgenic reporters and cell type specific antibodies, that somatic abducens motor neurons and a small subset of OPCs arise from common olig2+ neuroepithelial precursors in rhombomeres r5 and r6, but that all other motor neurons and OPCs do not similarly develop from shared pools of olig2+ precursors. In the absence of olig2 function, r5 and r6 precursors remain in the cell cycle and fail to produce abducens motor neurons, and OPCs are entirely lacking in the hindbrain. These studies, therefore, reveal both common and independent roles for olig2 in development of somatic motor neurons and oligodendrocytes of the hindbrain.
Introduction
In the spinal cord of vertebrate embryos, motor neurons and most oligodendrocytes, the myelinating cell type of the CNS, have a common origin. Dividing, neuroepithelial precursors that occupy ventral spinal cord and express the transcription factor Olig2 produce first motor neurons and then oligodendrocyte progenitor cells (OPCs), as well as some ventral interneurons, astrocytes and ependymal cells (Lu et al., 2000; Takebayashi et al., 2000; Zhou et al., 2000; Park et al., 2004; Masahira et al., 2006). These precursors, called pMN cells, are specified by graded distribution of the morphogen Sonic Hedgehog (Ericson et al., 1997; Briscoe and Ericson, 1999; Jessell, 2000), and Olig2 function is necessary for development of both motor neurons and OPCs (Lu et al., 2002; Park et al., 2002; Takebayashi et al., 2002; Zhou and Anderson, 2002). In mouse embryos deficient for Olig2 and the related Olig1 gene, pMN precursors instead produce V2 interneurons and astrocytes (Zhou and Anderson, 2002). Some OPCs also arise within dorsal spinal cord, and in dorsal hindbrain, independent of motor neuron origins (Cameron-Curry and Le Douarin, 1995; Sussman et al., 2000; Cai et al., 2005; Fogarty et al., 2005; Vallstedt et al., 2005).
The developmental relationship of motor neurons and oligodendrocytes in the hindbrain has not been investigated as thoroughly as in spinal cord. Hindbrain, or cranial, motor neurons form three classes, visceral, somatic and branchiomotor, based upon their synaptic targets and columnar organization, and develop as groups within segmentally iterated rhombomeres (r) (Chandrasekhar, 2004; Guthrie, 2007). Many hindbrain oligodendrocytes arise from ventral neuroepithelium (Ono et al., 1997; Davies and Miller, 2001; Vallstedt et al., 2005), but these cells may originate from discrete focal clusters rather than from a continuous longitudinal domain as in the spinal cord (Spassky et al., 1998; Perez Villegas et al., 1999; Miller, 2002). The correlation of these putative focal clusters to hindbrain motor neuron origins is not known. Notably, somatic cranial motor neurons fail to develop in Olig2 mutant mouse embryos, or in embryos lacking functions of Olig2 and the related Olig1 gene, whereas other motor neuron classes appear to form normally (Lu et al., 2002; Zhou and Anderson, 2002). Olig1;Olig2 mutant mice also lack all hindbrain oligodendrocytes (Zhou and Anderson, 2002). Whether hindbrain oligodendrocytes arise from Olig+ precursors that also produce somatic motor neurons, or somatic motor neurons and oligodendrocytes have independent requirements for Olig gene function has not been resolved.
Here, we report an investigation of hindbrain motor neuron and oligodendrocyte development using zebrafish as a model system. Through a combination of gene expression analysis, fate mapping and time-lapse imaging, we determined that abducens motor neurons and some oligodendrocytes arise from common olig2+ precursors in r5 and r6, but that all other hindbrain motor neurons and oligodendrocytes appear to develop independently of each other. In the absence of olig2 function, r5 and r6 olig2+ precursors are not specified for neuronal or glial fates but maintain their neuroepithelial characteristics.
Materials and Methods
Wild-type and transgenic zebrafish.
Wild-type and transgenic fish were raised in the Vanderbilt University Zebrafish Facility and embryos collected from pair matings. The embryos, raised at 28.5°C, were staged according to morphological criteria (Kimmel et al., 1995) and hours postfertilization (hpf). We used the following transgenic lines: Tg(olig2:egfp)vu12 (Shin et al., 2003), Tg(isl1:egfp) (Higashijima et al., 2000), Tg(olig2:DsRed2)vu19 (Kucenas et al., 2008) and Tg(nkx2.2a:megfp)vu17 (Kirby et al., 2006). The Tg(olig2:Kaede)VU85 transgenic line was produced using the same strategy that created the Tg(olig2:egfp) and Tg(olig2:DsRed2) lines (Shin et al., 2003; Kucenas et al., 2008).
In situ RNA hybridization.
The following previously described RNA probes were used: egr2b (also known as krox20) (Oxtoby and Jowett, 1993), isl1 (Inoue et al., 1994), sox10 (Dutton et al., 2001), olig2 (Park et al., 2002), and gfp. Embryos were fixed in 4% paraformaldehyde (PFA) and then stored in 100% methanol at −20°C. The in situ RNA hybridization was performed as previously described (Hauptmann and Gerster, 2000) followed by a color reaction with BM purple (Roche Diagnostics). For double RNA labeling probes were labeled with either digoxygenin or fluorescein. The first probe was detected with the appropriate antibody conjugated to alkaline phosphatase (AP) and followed by a color reaction with BM purple. Washing the embryos with 0.1 m glycine, pH 2.2 followed by a 20 min incubation with 4% PFA inactivated the first antibody and the appropriate second antibody was then applied and developed with a solution of INT/BCIP (Roche Diagnostics) (2-[4-iodophenyl]-3-[4-nitrophenyl]-5-phenyltetrazolium chloride)/(5-bromo-4-chloro-3-indolyl phosphate, toluidine salt in DMSO). Once developed, the embryos were dissected from the yolk and mounted in 70% glycerol for whole-mount imaging on bridged coverslips. All images were captured using Openlab software (Improvision) using an Olympus AX70 microscope equipped with DIC optics and a Retiga Exi 1300 color digital camera. Once captured, images were imported into Adobe Photoshop and adjustments were limited to contrast, levels, color matching settings and cropping.
Immunohistochemistry.
For immunohistochemistry we used the following primary antibodies: mouse anti-Isl [39.4D5, 1:100; Developmental Studies Hybridoma Bank (DSHB)], mouse anti-Zn8 (1:1000; DSHB), mouse anti-BrdU (G3G4, 1:200; DSHB), mouse anti-Pax7 (DSHB), rabbit anti-Sox10 (1:500) (Park et al., 2004), rabbit anti-Pax2 (1:100; Berkeley Antibody), mouse anti-Nkx6.1 (1:1000; DSHB), rabbit anti-GABA (1:1000; Millipore), mouse anti-GFAP (1:400; Sigma), mouse anti-Zrf1 (1:250; DSHB), mouse anti-HuC (1;100; Invitrogen) and rabbit anti-Calretinin (1:1000; Swant). For fluorescent detection, the following Alexa Fluro secondary antibodies were used: 568 goat anti-mouse, 568 goat anti-rabbit, 647 goat anti-mouse, and 647 goat anti-rabbit (all at 1:200; Invitrogen). Embryos were fixed in 4% AB fix (4% PFA, 8% sucrose, 1× PBS) for 2 h at room temperature (RT) or overnight at 4°C. Embryos for sectioning were embedded in 1.5% agar/5% sucrose, frozen with 2-methyl-butane chilled by immersion in liquid nitrogen, and sectioned using a cryostat microtome (10 μm). Sections were rehydrated with 1× PBS and preblocked for 30 min in 2% sheep serum/BSA-1× PBS. The sections were incubated with primary antibody overnight at 4°C, washed extensively with 1× PBS and incubated with the appropriate fluorescent secondary antibody for 2 h at RT. Once the secondary antibody was washed off sections were covered with Vectashield (Vector Laboratories).
Embryos for whole-mount imaging were fixed as above and preblocked with 10% sheep serum/BSA-1× PBS for 1 h at RT. The embryos were incubated in primary antibody for 24 h at 4°C, washed semicontinuously with 1× PBS for 2 h at RT, and then incubated with the secondary antibody for 12 h at 4°C, followed by 3 h of semicontinuous washes with 1× PBS. These embryos were then dissected from the yolk and mounted on bridged coverslips in 70% glycerol for imaging. All imaging was conducted on a Zeiss Axiovert 200 inverted microscope equipped with either a 40× oil-immersion objective [numerical aperture (NA) = 1.3] or 20× dry objective (NA = 0.75), mounted on a Piezo drive, and a PerkinElmer Ultraview ERS Live Cell Imager. Images were exported and analyzed using Volocity (Improvision) and Adobe Photoshop. Image adjustments were limited to level settings, contrast and cropping.
Bromodeoxyuridine labeling.
Embryos were manually dechorionated and incubated with 10 mm bromodeoxyuridine (BrdU) in 10% DMSO in embryo medium for 20 min on ice. For longer incubations, a 20 min pulse on ice was followed by incubation with 10 mm BrdU solution in embryo medium at 28.5°C. The embryos were then fixed using 4% pfa in PBS and sectioned as described above. Before anti-BrdU immunocytochemistry the sections were treated for 30 min with 2 m HCl. All sections were imaged using the Zeiss Axiovert 200 microscope described above.
Antisense morpholino oligonucleotide injections.
An antisense morpholino (MO) oligonucleotide with the sequence 5′-ACACTCGGCTCGTGTCAGAGTCCAT-3′ (Gene Tools) was designed to the olig2 translation start site. We also used a Standard Control MO (Gene Tools). Both morpholinos were resuspended in distilled water for a stock solution of 3 mm. The stock solution was further diluted with water and phenyl red and 1–2 ng was injected into the yolk of one- to two-cell stage embryos.
Conditional expression experiments.
The plasmid p(hsp70l:olig2)Tol2 was constructed by subcloning olig2 cDNA into a vector carrying the zebrafish hsp70l heat-responsive promoter (Shoji et al., 1998) and recognition sequences for Tol2 transposase (Kawakami, 2004). Newly fertilized eggs produced by Tg(olig2:egfp) or Tg(olig2:DsRed2) adults were injected with a solution containing 0.1 μg/μl of the plasmid and 0.3 μg/μl in vitro synthesized Tol2 mRNA. Expression was induced by transferring embryos at selected stages to egg water prewarmed to 39°C for 30 min.
Photoconversion.
Tg(olig2:Kaede) embryos were grown in embryo medium, containing 0.003% phenyl-thiourea (PTU) to prevent the formation of dark pigment, at 28.5C. They were kept in the dark to minimize photoconversion of the Kaede protein by ambient light. To photoconvert the Kaede protein, embryos were lightly anesthetized with 3-aminobenzoic acid ethyl ester (Tricaine) and immersed in 0.8% low-melting temperature agarose. They were then mounted in either lateral or dorsal orientations in glass-bottom 35 mm Petri dishes. Using a Zeiss Axioskop 2 FS microscope equipped with a nitrogen pulsed laser, 405 nm dye cell, yellow beamsplitter, 40× water-immersion lens and green fluorescent protein (GFP) filter, small clusters of Kaede+ cells were photoconverted with ∼4 s laser pulses. The live embryos were then imaged using the confocal microscope described above.
Time-lapse imaging.
The embryos were manually dechorionated at 24 hpf and transferred to embryo medium containing PTU. Embryos for time-lapse imaging were anesthetized using Tricaine and immersed in 0.8% low-melting temperature agarose. They were then mounted in either lateral or dorsal orientations in glass-bottom 35 mm Petri dishes. Images were captured using a 20× dry (NA = 0.75) objective mounted on the confocal microscope described above. A heated stage and chamber kept the embryos at 28.5°C and Z image stacks were collected every 5–15 min. The data sets were analyzed using Volocity software and exported as QuickTime files to create movies.
Results
olig2 expression reveals common precursors for abducens motor neurons and oligodendrocytes
We initiated our studies by using in situ hybridization to determine the distribution of olig2 RNA (Park et al., 2002) in the hindbrain of zebrafish embryos. olig2+ cells were first evident by 22 hpf at the level of the otic vesicle (data not shown). By 28 hpf olig2+ cells were located in two distinct bilateral clusters at the midline of the posterior hindbrain (Fig. 1A). Using egr2b (previously known as krox20) RNA expression as a marker for r3 and r5 (Oxtoby and Jowett, 1993), we determined that these olig2+ clusters occupy r5 and r6 (data not shown). By 40 hpf a third olig2+ population appeared just posterior to the original two (Fig. 1B). Subsequently, olig2 expression expanded anteriorly and posteriorly along the midline of the hindbrain (Fig. 1C,D) and by 3 d postfertilization (dpf) appeared throughout the hindbrain (data not shown).

RNA expression profiles of hindbrain motor neuron and OPC markers. All panels show dorsal views of whole embryos, anterior to the left. A–D, olig2 expression begins in r5 and r6 at the midline and extends posteriorly and then anteriorly. E–H, egfp expression in Tg(olig2:egfp) embryos is similar to olig2 expression. I–L, isl1 expression marks the cell bodies of cranial nerves V, VII, X at 28 hpf and cranial nerves VI and IX beginning at 40 hpf. isl1 also labels trigeminal ganglion cells (tg). M, Schematic of hindbrain motor neuron organization. Branchial motor neurons are depicted in red, somatic motor neurons in green. CHB, Caudal hindbrain. N–P, sox10 expression (arrow) begins at 40 hpf at the midline near the otic vesicle (ov) and extends along the anteroposterior axis by 48 and 58 hpf. sox10 expression also marks neural crest (nc) cells migrating over the surface of the hindbrain.
Enhanced GFP (EGFP) driven by olig2 regulatory DNA precisely recapitulates olig2 expression in the spinal cord (Shin et al., 2003). To compare reporter gene expression to endogenous olig2 expression in the hindbrain, we performed in situ hybridization to detect egfp RNA in Tg(olig2:egfp) transgenic embryos. Similar to olig2, egfp RNA was first evident in two distinct clusters in r5 and r6 and then along the anteroposterior length of the hindbrain (Fig. 1E–H). Therefore, as in the spinal cord, transgenic reporter gene expression is an accurate indicator of hindbrain olig2 expression.
To compare the distribution of olig2+ cells to the distribution of hindbrain motor neurons, we examined the expression of isl1 RNA, a motor neuron marker (Korzh et al., 1993). isl1 was first detected at 16 hpf (data not shown) in the anterior hindbrain. By 28 hpf distinct clusters of isl1+ cells were located along the anteroposterior axis, corresponding to cranial nerves V, VII, and X (Fig. 1I), as previously described (Chandrasekhar et al., 1997; Higashijima et al., 2000). Cranial nerves V and X arise and remain in the rhombomeres of their birth, r2/3 and caudal hindbrain respectively, whereas the cell bodies for cranial nerve VII are born in r4 and subsequently migrate into r6 and r7 (Chandrasekhar, 2004). At 40 hpf cranial nerve VI was present in r5 and r6 and we observed the cell bodies of cranial nerve VII along the midline as they migrated from r4 to r6 and r7 (Fig. 1J). Migration was completed soon after and the final pattern of hindbrain motor neurons, revealed by isl1 expression, was evident at 48 and 58 hpf (Fig. 1K–M).
We also used in situ RNA hybridization to detect the expression of sox10, which specifically marks oligodendrocyte lineage cells in the CNS, including oligodendrocyte progenitor cells (OPCs), dividing and migratory cells that give rise to myelinating oligodendrocytes (Kuhlbrodt et al., 1998). At 28 hpf, no sox10 expression was evident indicating that, like spinal cord, OPCs are produced after motor neurons (data not shown). By 40 hpf, a small cluster of sox10+ cells was apparent at about the position of r5 (Fig. 1N). Subsequently, sox10 expression expanded anteriorly and posteriorly similar to olig2 expression (Fig. 1O,P).
The correlation of olig2, isl1 and sox10 expression in r5 and r6 raised the possibility that these rhombomeres are sites of common precursors for oligodendrocytes and motor neurons. As a first test of this possibility, we examined the morphology of olig2+ cells marked by EGFP expression in Tg(olig2:egfp) embryos. EGFP fluorescence was evident by 33 hpf in two prominent clusters occupying r5 and r6 (Fig. 2A). This pattern is similar to that revealed by in situ RNA hybridization, but somewhat later, reflecting the time necessary to translate and fold EGFP. Transverse and sagittal sections revealed that olig2+ cells were ventral and medial within the hindbrain and extended fine processes to the pial surface, characteristic of neuroepithelial cells (Fig. 2E,I). By 48 hpf, olig2+ cells appeared at the hindbrain midline both anterior and posterior to the r5 and r6 clusters and some cells with elongated morphologies occupied more lateral positions (Fig. 2B). High magnification views of sections showed some cells within the r5 and r6 clusters with processes extending to the pial surface and others with thicker processes that exited the hindbrain ventrally and turned anteriorly, suggestive of axonal projections (Fig. 2F,J). Other cells, located outside the clusters, had very fine processes characteristic of OPCs. By 58 hpf, the r5 and r6 clusters were less distinct and more olig2+ cells occupied lateral positions (Fig. 2C). Fewer neuroepithelial cells were evident within the clusters but cells with ventrally and anteriorly extending processes were still present (Fig. 2G,K). By 3 dpf, the r5 and r6 olig2+ clusters were no longer visible but individual cells were dispersed throughout the hindbrain (Fig. 2D). High magnification views showed that olig2+ cells had OPC-like morphologies, extending numerous fine processes to form a dense meshwork (Fig. 2H,L).

Morphologies of hindbrain olig2+ cells revealed by transgenic reporter expression. All images show EGFP fluorescence in Tg(olig2:egfp) embryos. A–D, Dorsal views of whole embryos, anterior to the left. r5 and r6 indicate EGFP expression at the midline in r5 and r6. Dashed lines mark edges of neural tube. Scale bar, 48 μm. E–H, Transverse sections through the hindbrain, dorsal up, either 30 μm (F) or 10 μm (E, G, H) thick. At 33 hpf (E), EGFP+ cell bodies are clustered at the midline and extend processes to the pial surface. From 48 hpf onward (F–H), cells with fine membrane protrusions characteristic of OPCs are present (arrows). Arrowheads indicate thick, ventrally extending processes. Scale bar, 24 μm. I–L, Sagittal sections, dorsal up, anterior to the left, either 30 μm (J) or 10 μm (I, K, L) thick. Dotted lines mark the edge of the neural tube. Open arrowheads indicate axon-like extensions that exit the hindbrain ventrally and turn anteriorly. Scale bar, 24 μm.
The static images presented above show that olig2+ cells are first at the ventral medial neural tube and then occupy the entire hindbrain. To better understand how olig2+ cells become distributed, we performed time-lapse imaging. This revealed that some olig2+ cells with OPC morphologies migrated from the r5 and r6 clusters and confirmed that other cells within the clusters were the source of apparent axons that extended ventrally and then anteriorly (supplemental Movie 1, available at www.jneurosci.org as supplemental material). Additionally, many other olig2+ OPC-like cells emerged individually along the anteroposterior axis of the hindbrain, both within the ventral and dorsal neural tube, and subsequently migrated along the dorsoventral and anteroposterior axes (supplemental Movies 1 and 2, available at www.jneurosci.org as supplemental material). These observations suggested that olig2+ neuroepithelial precursors in r5 and r6 produce some OPCs and motor neurons but that many OPCs arise from olig2− precursors throughout the remainder of the hindbrain.
By morphology, hindbrain olig2+ cells appeared to include both motor neurons and OPCs. To obtain supporting evidence and determine the motor neuron identities we labeled Tg(olig2:egfp) embryos with various cell type specific markers. At 40 hpf, no cells were labeled with anti-Sox10 antibody (Park et al., 2005) indicating that OPCs were not yet specified (Fig. 3A). Sox10+ cells appeared by 48 hpf, and each cell was also olig2+ (Fig. 3B). All olig2+ multiprocess cells outside the r5 and r6 clusters were Sox10+, confirming their identity as OPCs. Notably, some r5 and r6 cluster cells also expressed Sox10 (Fig. 3B), consistent with the possibility that neuroepithelial cells within the clusters are precursors for OPCs. By 64 hpf, nearly every olig2+ cell in the hindbrain was Sox10+, with only a few cells with long, ventrally extending processes remaining Sox10− (Fig. 3C). By 3 dpf, olig2 and Sox10 expression completely coincided (Fig. 3D) indicating that all hindbrain Sox10+ OPCs express olig2.

OPCs and abducens motor neurons express olig2. A–D, Anti-Sox10 immunohistochemistry to mark OPCs in sagittal (A, B) and transverse (C, D) sections of Tg:(olig2:egfp) embryos. Arrows and arrowheads mark olig2+ Sox10+ cells outside and within olig2+ r5 and r6 clusters, respectively. Open arrowheads mark axonal projections. E–H, Anti-Isl antibody immunohistochemistry, shown in red, to mark motor neurons in sagittal (E–G) and transverse (H) sections of Tg:(olig2:egfp) embryos. Arrows mark olig2+ Isl+ motor neurons and open arrowhead points to axonal projections. I, J, Sagittal sections of Tg(isl1:egfp); Tg(olig2:DsRed2) embryos. The transgenic reporters mark distinct cell populations. K–P, Zn8 immunohistochemistry, shown in red, to mark abducens motor neurons in Tg:(olig2:egfp) embryos. Whole embryos viewed from dorsal, anterior to the left (K, L) and transverse (N, P) and sagittal sections (O), dorsal to the top. Arrows mark olig2+ Zn8+ abducens motor neurons and projections. Dashed lines indicate neural tube boundary. Scale bars: (A–J, N–P) 24 μm; (K–M) 48 μm.
We next used anti-Isl antibody to label all motor neuron cell bodies in the hindbrain of Tg(olig2:egfp) embryos (Chandrasekhar et al., 1997). Whereas no olig2+ cells expressed Isl at 33 hpf, numerous cells within the r5 and r6 olig2+ clusters were labeled by anti-Isl antibody at 48 hpf (Fig. 3E,F). However, many Isl+ cells in r5 and r6 did not express olig2 at 48 hpf and 56 hpf (Fig. 3F,G), nor did any other Isl+ motor neurons in midbrain or hindbrain express olig2 between 24 hpf and 3 dpf (supplemental Fig. 1A–D, available at www.jneurosci.org as supplemental material), consistent with the observation that, in mice, Olig2 function is necessary for development of only a subset of motor neurons (Lu et al., 2002; Zhou and Anderson, 2002; Gaufo et al., 2003; Pattyn et al., 2003). By 3 dpf double labeling was no longer evident (Fig. 3H), suggesting that, as in the spinal cord, motor neurons downregulate olig2 expression.
Isl protein is localized to nuclei and so anti-Isl labeling does not reveal other features of cell identity except for position within the neural tube. Consequently, to more definitively identify olig2+ motor neurons, we examined Tg(isl1:egfp); Tg(olig2:DsRed2) transgenic embryos. EGFP expressed by the isl1 transgene is cytoplasmic, permitting visualization of motor neuron cell bodies and their axons (Higashijima et al., 2000). Although this particular transgenic reporter labels most cranial motor neurons, it does not label abducens motor neurons (Higashijima et al., 2000), which are located in r5 and r6 in zebrafish (Moens et al., 1996). In contrast to our anti-Isl antibody labeling results, at 48 hpf and 58 hpf the transgenes were not expressed in the same cells (Fig. 3I–J). This raised the possibility that the r5 and r6 olig2+ motor neurons are abducens motor neurons. To test this, we next used Zn8 antibody, which recognizes the cell surface protein Neurolin, also known as Dm-Grasp (Kanki et al., 1994; Chandrasekhar et al., 1997). Zn8 did not label any olig2+ cells at 32 hpf, however, by 36 hpf some double labeling was evident (Fig. 3K,N). At 48 and 60 hpf, Zn8 clearly labeled a subset of olig2+ cells, including their ventral projections, within the r5 and r6 clusters (Fig. 3L,O,P). Consistent with the Isl antibody labeling, very little double labeling with Zn8 persisted to 3 dpf (Fig. 3M,P). Therefore, olig2+ r5 and r6 cells include abducens motor neurons. Altogether, these data indicate that, in the zebrafish hindbrain, olig2 expression marks all OPCs but only abducens motor neurons.
The close proximity of olig2+ abducens motor neurons and some OPCs in r5 and r6 raised the possibility that they arise from common populations of dividing, olig2+ neuroepithelial precursors. To test this, we treated Tg(olig2:egfp) embryos with the thymidine analog BrdU to label cells in S phase. olig2+ BrdU+ cells were evident in r5 and r6 at 30 and 33 hpf, but by 48 hpf significantly fewer cells incorporated BrdU throughout the hindbrain (Fig. 4A–C), reflecting a dramatic reduction in the dividing cell population as previously reported (Lyons et al., 2003). By 56 hpf, we found no olig2+ BrdU+ cells in ventromedial r5 and r6, which coincides with the loss of olig2+ cells with neuroepithelial morphology noted above. Instead, the only olig2+ cells labeled by BrdU at this time were OPCs (Fig. 4D). These data show that olig2+ r5 and r6 cells divide, consistent with the possibility that these cells are precursors for both abducens motor neurons and OPCs.

olig2+ neuroepithelial cells and OPCs are mitotically active. Anti-BrdU immunohistochemistry, shown in red, to reveal S-phase cells in Tg(olig2:egfp) embryos. A–C, Transverse sections obtained from 30, 33 and 48 hpf embryos reveal olig2+ BrdU+ neuroepithelial cells (arrows). D, Sagittal section. By 56 hpf, olig2+ neuroepithelial cells are no longer evident and only OPCs incorporate BrdU (arrows). Dotted lines mark the edge of the neural tube. Scale bar, 24 μm.
We next used fate mapping to directly investigate the fates of olig2+ r5 and r6 cells. To do so we created a Tg(olig2:Kaede) transgenic line, which expresses the Kaede photoconvertable fluorescent protein under the control of olig2 regulatory DNA. As expected, photoconversion of either r5 or r6 clusters labeled cells with axonal projections characteristic of abducens motor neurons (Fig. 5A–H). Additionally, by 60 hpf a small number of photoconverted cells with OPC morphologies were outside of the original clusters (Fig. 5C,D,G,H). Notably, many OPCs expressed only the green form of Kaede, indicating that they did not arise from a photoconverted cluster, even when both clusters were photoconverted (data not shown). Therefore, r5 and r6 olig2+ precursors produce abducens motor neurons and a subset of hindbrain OPCs.

Fate mapping reveals that abducens motor neurons and some OPCs arise from r5 and r6 olig2+ cells. A–D and E–H show images from two different Tg(olig2:Kaede) embryos. In both cases, olig2+ cells where photoconverted at 40 hpf and imaged at 50 hpf and 60 hpf. D and H show only the red channel, revealing the photoconverted cells. Arrows indicate photoconverted cells that migrated from their origins and arrowheads mark abducens motor axon projections. Scale bar, 48 μm.
olig2 is required for OPC and abducens motor neuron specification
To determine the requirement of olig2 in OPC and motor neuron specification in the hindbrain, we injected into newly fertilized Tg(olig2:egfp) eggs an olig2 antisense morpholino (MO) designed to block translation of endogenous olig2 transcripts but not of egfp mRNA driven by the transgene. This allowed us to assess the fates of cells that express olig2 in the absence of olig2 function. As we showed previously (Park et al., 2002), injected embryos produced few spinal cord motor neurons and oligodendrocytes (data not shown). 33 and 48 hpf MO-injected embryos had few olig2+ cells in the hindbrain outside of the r5 and r6 clusters (Fig. 6A–D), suggesting that they had a deficit of OPCs. The r5 and r6 olig2+ clusters were present in MO-injected embryos, however, most of the cells appeared to have neuroepithelial morphologies, and axonal morphologies characteristic of abducens motor neurons were not evident (Fig. 6E–H). Labeling MO-injected embryos with anti-Sox10 antibody confirmed the near absence of hindbrain OPCs, both within and outside of the r5/r6 olig2+ clusters (Figs. 6I–L; 7A). Although the general pattern of all other Isl+ cranial motor neurons appeared unaffected, including the anterior somatic motor neurons (Fig. 6M–P; supplemental Fig. 1E,F, available at www.jneurosci.org as supplemental material), MO-injected embryos had few Isl+ olig2+ cells in r5 and r6, indicating the absence of only abducens motor neurons. Consistent with this, MO-injected embryos also had a deficit of Zn8+ olig2+ r5 and r6 cells between 48 and 56 hpf (Figs. 6Q–X; 7B). To further confirm the loss of OPCs and abducens motor neurons in MO-injected embryos, we used the Tg(olig2:Kaede) line to determine the fate of olig2+ cells. Consistent with the antibody labeling, we found an absence of photoconverted cells outside of the original r5/r6 clusters and none of the photoconverted cells had axonal projections characteristic of abducens motor neurons (supplemental Fig. 2, available at www.jneurosci.org as supplemental material). Therefore, olig2 is required for specification of abducens motor neurons, in addition to OPCs.

OPC and abducens motor neuron development require olig2 function. All panels show Tg(olig2:egfp) embryos. A–D, Dorsal views of whole wild-type (A,C) and olig2 MO-injected (B, D) embryos, anterior to the left. Wild-type and MO-injected embryos appear similar at 33 hpf (A, B) but at 48 hpf MO-injected embryos have few dispersed OPCs compared with wild type (C, D). E, F, Transverse sections showing that olig2+ neuroepithelial cells appear similar in wild-type and MO-injected embryos at 33 hpf. G, H, Sagittal section of 56 hpf embryos. No olig2+ cells with OPC and motor neuron morphologies are apparent in the MO-injected embryo. Open arrowhead marks motor axon projections in wild type. I–L, Anti-Sox10 immunohistochemistry (blue) at 48 hpf (I, J, sagittal sections) and 56 hpf (K, L; transverse sections) reveals absence of OPCs in MO-injected embryos. Arrows point to olig2+ Sox10+ OPCs in wild type. M–P, Anti-Isl immunohistochemistry (red) at 48 hpf (M, N; dorsal view, whole embryos) and 56 hpf (O, P; sagittal sections). The general pattern of Isl+ cranial motor neurons is unaffected in MO-injected embryos (M, N) but MO-injected embryos have fewer olig2+ Isl+ motor neurons than wild type (N, P). Q–X, Zn8 immunohistochemistry (red) shown in transverse sections (Q, R), sagittal sections (S, T) and whole embryos from dorsal view (U–X). MO-injected embryos have a deficit of Zn8+ abducens motor neurons (arrows). Dotted lines mark the edge of the neural tube. Scale bars: (A–D, M–P, U–X) 48 μm; (E–L, Q–T) 24 μm.

Quantification of OPCs and abducens motor neurons. A, Average number of olig2+ Sox10+ OPCs per transverse section in wild-type, control MO-injected and olig2 MO-injected embryos. B, Average number of olig2+ Zn8+ abducens motor neurons per transverse section in wild-type, control MO-injected and olig2 MO-injected embryos. Statistical significance was determined using the unpaired t test. p values are shown for each treatment compared with wild type. Error bars represent the SEM.
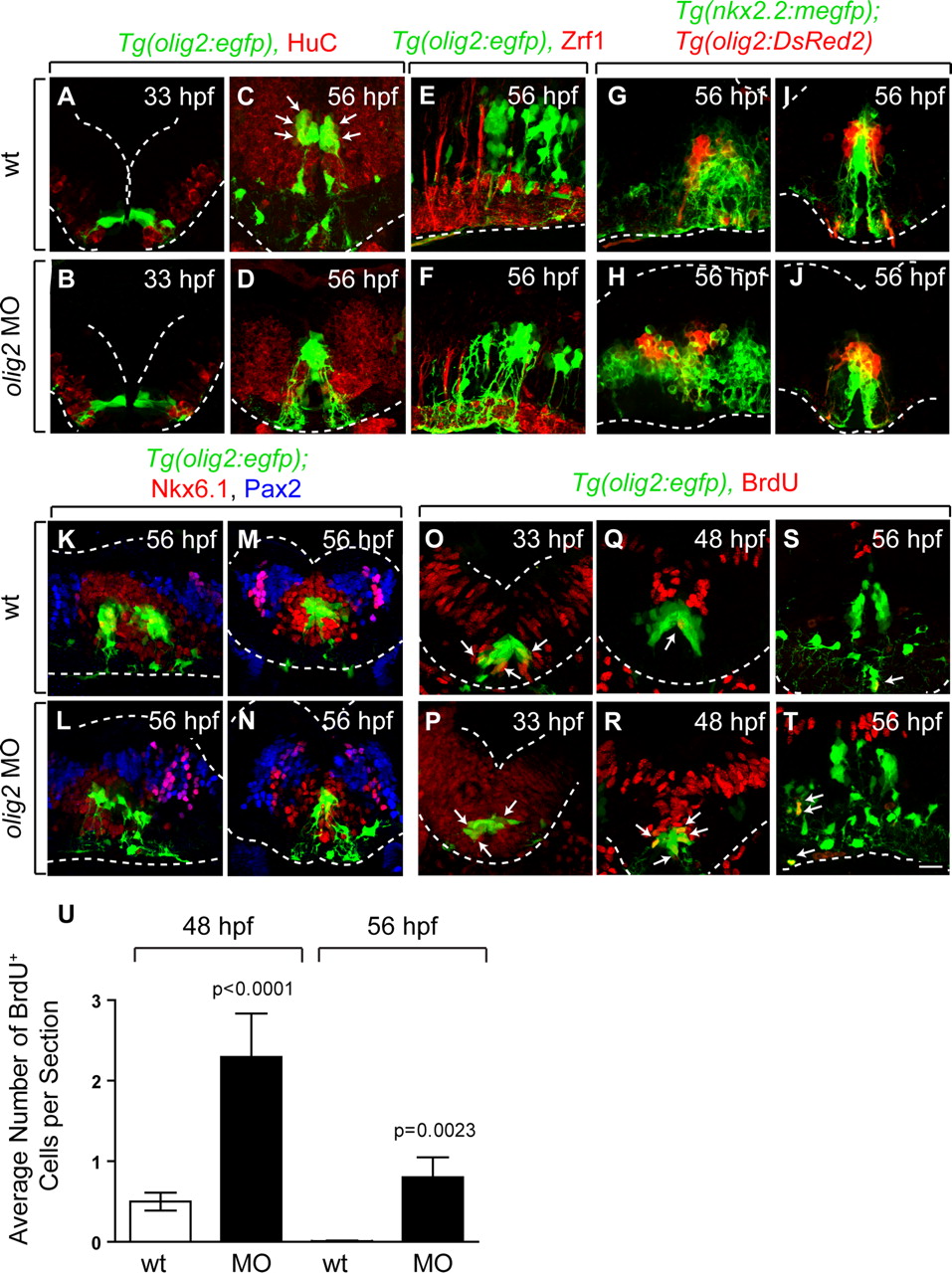
In mice, mutation of Olig genes alters dorsoventral patterning of the spinal cord so that cells that would normally have pMN precursor identity produce v2 interneurons and astrocytes instead of motor neurons and oligodendrocytes (Lu et al., 2002; Zhou and Anderson, 2002). Therefore, we sought to determine whether r5 and r6 olig2+ precursors are similarly specified for alternative neuronal and glial fates in the absence of olig2 function. To test this, we first labeled embryos with anti-HuC antibody, which marks all newly specified neurons (Marusich et al., 1994). Consistent with anti-Isl labeling, a subset of r5 and r6 olig2+ cells in wild-type embryos was HuC+ at 56 hpf but not at 33 hpf or 3 dpf (Fig. 8A,C) (data not shown). In contrast, MO-injected embryos had very few r5 and r6 olig2+ HuC+ cells at any stage (Fig. 8B,D) (data not shown). Next, we labeled wild-type and MO-injected embryos with Zrf1 antibody, which marks radial glia (Trevarrow et al., 1990) and GFAP antibody, which marks radial glia and astrocytes (Marcus and Easter, 1995). Neither antibody labeled olig2+ cells in wild-type or MO-injected embryos (Fig. 8E,F) (data not shown). Therefore, in the absence of olig2 function, r5 and r6 olig2+ precursors fail to differentiate as neurons or glial cells.

olig2+ cells retain neuroepithelial precursor characteristics in the absence of olig2 function. A–D, Anti-Hu immunocytochemistry (red) to reveal neurons in transverse sections. Wild-type and MO-injected embryos are similar at 33 hpf (A, B). At 56 hpf, MO-injected embryos have few olig2+ Hu+ neurons compared with wild type (C, D). E, F, Zrf1 immunohistochemistry (red) to reveal radial glia in sagittal sections. G–J, Sagittal (G, H) and transverse (I, J) sections of Tg(olig2:DsRed2);Tg(nkx2.2a:megfp) embryos. The general pattern of nkx2.2a expression (green) is unchanged in MO-injected embryos. K–N, Anti-Nkx6.1 (red) and anti-Pax2 (blue) immunocytochemistry on sagittal (K, L) and transverse (M, N) sections. O–T, Anti-BrdU immunohistochemistry (red) to reveal cells in S-phase. MO-injected embryos have more olig2+ BrdU+ neuroepithelial cells (arrows) at 48 and 56 hpf than wild type. U, Average number of olig2+ BrdU+ cells per transverse section in wild-type and olig2 MO-injected embryos. Statistical significance was determined using the unpaired t test. p values are shown for each treatment compared with wild type. Error bars represent SEM. Dotted lines mark the edge of the neural tube. Scale bar, 24 μm.
We further investigated hindbrain patterning using antibodies that recognize Pax2 and Nkx6.1, and the transgenic reporter Tg(nkx2.2a:megfp) as markers of hindbrain cell populations. At 56 hpf, nkx2.2a reporter expression marks ventral hindbrain cells that border olig2+ cells in wild-type embryos (Fig. 8G,I). Nkx6.1+ cells surround olig2+ cells and, in turn, are surrounded by Pax2+ cells (Fig. 8K,M). Although hindbrain patterning appears slightly perturbed in olig2 MO-injected embryos, the relationship of these gene expression patterns to one another remained unchanged (Fig. 8G–N). At 4 dpf, neuroepithelial olig2+ cells persisted at the r5/r6 midline in MO-injected embryos, whereas they were absent from wild type, but dorsoventral patterning, revealed by Nkx6.1, Pax2 and Pax7 expression, remained normal (supplemental Fig. 3, available at www.jneurosci.org as supplemental material).
The failure of r5 and r6 olig2+ hindbrain cells to produce neurons and glial cells in the absence of olig2 function raised the possibility that they remain as precursor cells. To test this we treated wild-type and MO-injected embryos with BrdU to mark cells in S-phase. At 33 hpf, there was no difference in the number of olig2+ r5 and r6 cells that incorporated BrdU in wild-type and MO-injected embryos (Fig. 8O,P). In contrast, whereas the number of olig2+ cells that incorporated BrdU in wild type declined by 48 hpf and 56 hpf, the number of olig2+ S-phase cells remained high in MO-injected embryos (Fig. 8Q–U). These data indicate that r5 and r6 neuroepithelial precursors require olig2 function to exit the cell cycle and differentiate.
In chick spinal cord, ectopic expression of Olig2 in combination with Nkx2.2, but not alone, can promote premature and ectopic formation of oligodendrocytes (Zhou et al., 2001). Additionally, ectopic expression of Olig2 in chick and zebrafish spinal cord causes dorsal expansion of the motor neuron population (Mizuguchi et al., 2001; Novitch et al., 2001; Park et al., 2002). To test the capacity of olig2 to promote formation of oligodendrocytes and motor neurons in the zebrafish hindbrain, we injected newly fertilized eggs with a plasmid that permits time-dependent expression of olig2 controlled by a heat-responsive promoter. We induced expression in separate sets of embryos at 16 and 24 hpf, fixed them either 2, 12 or 16 h later and performed Sox10 and Isl immunocytochemistry to detect OPCs and motor neurons, respectively. From a total of 66 experimental embryos, we never observed prematurely or ectopically produced OPCs or motor neurons (data not shown). Therefore, as in the spinal cord, olig2 is necessary, but apparently not sufficient, for OPC and motor neuron specification in the hindbrain.
Discussion
olig2 expression identifies neural precursors that produce abducens motor neurons and a subset of hindbrain oligodendrocytes
The best characterized source of OPCs are pMN precursors, which occupy ventral spinal cord, express Olig genes and produce motor neurons, some interneurons, astrocytes and ependymal cells in addition to OPCs (Novitch et al., 2001; Lu et al., 2002; Zhou and Anderson, 2002; Masahira et al., 2006). Subsequently, OPCs migrate radially, dorsoventrally and longitudinally to reach their target axons, whereupon they differentiate as mature oligodendrocytes. A smaller number of oligodendrocytes also originates from dorsal spinal cord, indicating that neither Olig gene expression by neuroepithelial precursors nor shared lineage with motor neurons is obligatory for OPC formation (Sussman et al., 2000; Cai et al., 2005; Fogarty et al., 2005; Vallstedt et al., 2005).
The origins of hindbrain OPCs have not been as clearly defined as in spinal cord. Expression of the oligodendrocyte lineage cell marker plp/dm20 suggested that OPCs arise from segmentally iterated domains along the hindbrain ventral ventricular zone of chick and mouse embryos, reflecting a rhombomeric organization (Timsit et al., 1995; Perez Villegas et al., 1999; Le Bras et al., 2005). Labeling of chick embryos with the O4 antibody, which also marks oligodendrocyte lineage cells (Orentas and Miller, 1996; Ono et al., 1997) likewise implicated a ventral ventricular zone origin of oligodendrocytes within the hindbrain as well as other more lateral and dorsal origins (Davies and Miller, 2001). At E13.5, mouse embryos express Olig1 and Olig2 within a ventral domain that extends along the entire anteroposterior axis of the hindbrain, with the exception of r1, and within the more dorsal alar plate (Vallstedt et al., 2005). Both ventral and dorsal hindbrain explants produce oligodendrocytes, providing strong evidence that hindbrain oligodendrocytes have multiple origins (Davies and Miller, 2001; Vallstedt et al., 2005).
Whereas spinal motor neurons arise from pMN precursors to form columns along the length of the cord, cranial motor neurons have different dorsoventral origins and are organized as discrete nuclei within the midbrain and hindbrain (Chandrasekhar, 2004; Guthrie, 2007). Like spinal motor neurons, somatic cranial motor neurons emerge from the pMN precursor domain but branchiomotor and visceral cranial motor neurons originate from more ventral p3 precursors (Pattyn et al., 2003). Somatic motor neuron nuclei consist of three that innervate eye muscles, oculomotor, trochlear and abducens, and one, hypoglossal, that innervates tongue muscles (Guidato et al., 2003). The oculomotor and trochlear motor neurons occupy midbrain and r1, respectively, and abducens motor neurons are located in r5 of mice and r5 and r6 of chick and zebrafish (Moens and Prince, 2002; Guidato et al., 2003; Guthrie, 2007). Hypoglossal motor neurons form within r8 of mouse and chick but may be absent from zebrafish (Chandrasekhar, 2004).
These characterizations of oligodendrocyte and motor neuron origins reveal a potentially interesting difference between spinal cord and hindbrain. In the spinal cord apparently all spinal cord pMN precursors produce both OPCs and motor neurons (Lu et al., 2002; Park et al., 2002; Zhou and Anderson, 2002; Park et al., 2004; Masahira et al., 2006). In the hindbrain, some pMN precursors, for example those in r5 and r7 in mouse, might similarly produce somatic motor neurons and OPCs, whereas others might give rise only to OPCs. Our investigation of cranial motor neuron and hindbrain OPC development in zebrafish now provides evidence for this possibility. In the spinal cord, expression of olig2 appears first in an uninterrupted column of dividing, neuroepithelial precursors and is then maintained by descendent OPCs and oligodendrocytes. In contrast, in the zebrafish hindbrain olig2 RNA is first evident within two clusters of cells in r5 and r6. Only later do more anterior and posterior hindbrain cells begin to express olig2. EGFP expression driven by an olig2 transgene revealed differences in the morphologies of cells along the anteroposterior hindbrain axis as they initiated olig2 expression. Only the cells that expressed olig2 earliest in r5 and r6 had morphologies characteristic of neuroepithelial cells, which we confirmed by showing that they incorporate BrdU. EGFP+ cells in all other rhombomeres had morphologies and migratory behaviors characteristic of OPCs. Examination of transgenic reporter gene expression and labeling by cell-type-specific antibodies revealed that, of the different cranial motor neuron classes only abducens motor neurons express olig2. The close association of abducens motor neurons and some OPCs to olig2+ neuroepithelial cells in r5 and r6 suggested that the latter serve as multipotent precursors. The results of our fate mapping experiments, using photoconversion of a transgenic reporter, are consistent with this although it is important to note that because we did not label single cells we do not know if the same neuroepithelial precursor can produce motor neurons and OPCs as they do in the spinal cord (Park et al., 2004). In all other regions of the hindbrain, both ventral and dorsal, olig2 expression is initiated in OPCs and not the neuroepithelial precursors from which they arise. Therefore, our results indicate that the timing of olig2 expression within a cell lineage correlates with cell fate potential. Among hindbrain neuroepithelial precursors, those that express olig2 produce both somatic motor neurons and oligodendrocytes whereas those that do not express olig2 give rise to OPCs but not somatic motor neurons. This raises the possibility that differences in expression of Olig2 genes underlie species-specific distributions of somatic motor neurons.
olig2 is required for cell cycle exit and specification of motor neuron–OPC precursors
Olig1/Olig2 gene functions have been shown previously to be necessary for development of oligodendrocytes and motor neurons, including hindbrain somatic motor neurons (Lu et al., 2002; Zhou and Anderson, 2002; Gaufo et al., 2003; Pattyn et al., 2003). In the spinal cord of mouse embryos lacking both gene functions, pMN precursors give rise to V2 interneurons and astrocytes rather than motor neurons and oligodendrocytes (Zhou and Anderson, 2002) suggesting that pMN precursors take the fate of more dorsal p2 precursors. In contrast, we found that in zebrafish embryos lacking olig2 function, r5 and r6 precursors do not adopt alternative neuronal or glial fates, but instead continue to divide and maintain their neuroepithelial characteristics. Notably, overexpression of Olig2 in chick reduced the number of spinal cord cells that incorporated BrdU, and increased the number that expressed the p27 cyclin dependent kinase inhibitor and a pan-neuronal marker (Novitch et al., 2001). Additionally, Olig1−/−;Olig2−/− mouse embryos had BrdU+ cells located outside the ventricular zone suggesting that pMN cells failed to exit the cell cycle before their migration to the marginal zone (Zhou and Anderson, 2002). Therefore, in both spinal cord and hindbrain Olig gene expression in neuroepithelial precursors promotes cell cycle exit and neurogenesis. This may occur through Ngn2, because pMN cells in Olig1−/−;Olig2−/− mouse embryos lack Ngn2 expression (Zhou and Anderson, 2002), over-expression of Olig2 promotes ectopic Ngn2 expression (Novitch et al., 2001) and Ngn2 promotes cell cycle exit and neurogenesis (Farah et al., 2000; Novitch et al., 2001). Formation of OPCs must then require that Ngn2 function is repressed within a subset of Olig+ precursors. One likely mechanism is signaling mediated by Notch receptors, because Notch activity is continuously required during development for specification of OPCs from pMN precursors (Park and Appel, 2003; Kim et al., 2008).
Olig2 function is required for formation of all oligodendrocytes, whether they arise from Olig+ or Olig− precursors. This suggests that, in addition to the role of Olig2 in forming the pMN precursor domain and promoting the transition of dividing precursors to postmitotic motor neurons, it has an independent function in promoting differentiation of oligodendrocyte lineage cells. Consistent with this, over-expression studies in chick indicated that Olig2 can work with the zinc finger transcription factor Zfp488 to promote precocious Mbp expression (Wang et al., 2006). The specific roles of Olig2 genes in neural development are, therefore, likely to be determined both by transcriptional control elements that independently direct Olig2 expression in neural precursors and OPCs and cell type specific cofactors that determine the regulatory targets of Olig2 proteins.
Footnotes
-
This work was supported by National Institutes of Health Grant NS046668 and Training in Fundamental Neuroscience Grant T32 MH064913 (D.A.Z.). We thank Jennifer Miller for creating the Tg(olig2:Kaede) transgenic line and members of the Appel laboratory for discussions. The anti-BrdU antibody, developed by S. J. Kaufman, anti-Isl antibody, developed by T. M. Jessell, anti-Nkx6.1 antibody, developed by O. D. Madsen, anti-Pax7 antibody, developed by A. Kawakami, and Zrf1 and Zn8 antibodies, developed by B. Trevarrow, were obtained from the Developmental Studies Hybridoma Bank developed under the auspices of the National Institute of Child Health and Human Development and maintained by The University of Iowa, Department of Biological Sciences, Iowa City, IA. Confocal microscopy was performed using equipment provided by the Vanderbilt Academic Venture Capital Fund and The Light Microscopy Core Facility at the University of Colorado, Anschutz Medical Campus.
- Correspondence should be addressed to Bruce Appel, Department of Pediatrics, University of Colorado Denver, Anschutz Medical Campus, Mail Stop 8108, Aurora, CO 80045. Bruce.Appel{at}uchsc.edu





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}