

Abstract
Cells can sense and interpret mechanical stimuli from their environment, but the ability to engineer customized mechanosensing capabilities has remained a synthetic biology challenge. Here, we introduce a set of synthetic Notch (SynNotch)-derived proteins that can be used to convert extracellular tensile forces into specifiable gene expression changes. By elevating the tension levels needed to induce SynNotch activation, in combination with structure-guided mutagenesis, we designed tunable mechanoreceptors with sensitivities in the physiologically relevant picoNewton (pN) range. Cells expressing these receptors could distinguish between varying levels of ligand-mediated tension and enact customizable transcriptional responses in turn. The utility of these tools was demonstrated by the design of a decision-making circuit, through which fibroblasts could be made to differentiate into myoblasts in response to mechanostimulation with tensile forces of distinct magnitudes. This work provides insight regarding how mechanically-induced structural alterations in proteins can be used to transduce physical forces into biochemical signals, and the system should facilitate further programming of force-related phenomena in biological systems.
Main Text
Mechanical forces are fundamental regulators of biology, guiding vital processes and shaping disease progressions that span from the molecular to the tissue scale (1–3). Mechanical forces can do so by transmitting actionable information from biological environments, which can vary throughout developmental and tissue contexts (4). Cells have evolved complex mechanisms to actively utilize mechanical information, and to translate force-based stimuli into biochemical responses that can affect changes in metabolism, migration, differentiation, immunity, and gene transcription (5–9). Harnessing cells’ capability to interpret differential forces in their environment would enable advances in tissue engineering, where forces could be used to drive structural organization, and novel targeted therapeutics, where mechanical environments could be used as tractable disease biomarkers. However, it has been difficult to synthetically reproduce mechanosensation in cells, as its underlying mechanisms remain incompletely elucidated.
At a fundamental level, mechanotransduction can be generalized as input-output relationships, in which the mechanical energy is converted into a biochemical responses through force-induced changes in the structure of cell signaling molecules. The sensitivity of proteins to mechanical unfolding has been tuned through several strategies, but in the context of in vitro spectroscopy techniques or FRET sensors rather than customized cellular signaling networks (10–16). Conversely, outputs from endogenous pathways have been modulated by overexpressing protein components, engineering sensitivity to non-mechanical inputs, and, more synthetically, adding novel outputs to the force-sensitive Piezo1 and YAP/TAZ signaling cascades (17–19). However, these approaches have not yet tuned the mechanical force required for activation, and do not present an obvious protein engineering solution for doing so. These strategies furthermore rely upon endogenous pathways that are particularly difficult to decouple from the user’s engineered pathways. Ideally, a fully customizable mechanosensitive pathway would synthesize several design criteria: programmable sensitivity to input forces, versatility of output cellular actions, and orthogonality to endogenous signaling pathways.
A simple platform for engineering cellular mechanosensation
Here, we use the Notch1 receptor to build a platform for controlling mechanosensation that meets the above design criteria. Notch1 is a classically studied protein, and our work capitalizes on a body of structural and biophysical research that guides our molecular design. Notch1 activation requires mechanical force, which is sensed by the receptor’s negative regulatory region (NRR) (Fig. 1A) (20). Upon application of tensile force via the ligand binding domain (LBD), the NRR undergoes a conformational change that enables proteolytic release of the intracellular domain (ICD), a transcriptional effector (20–23). Recently, work has shown that the NRR can act as a modular scaffold to build synthetic Notch (SynNotch) receptors, which preserve the Notch activation mechanism while tolerating heterologous substitutions at the LBD and ICD (24). These receptors offer a starting point satisfying two of our design criteria; SynNotch receptors provide versatility of cellular outputs via an ICD of interest and minimal crosstalk with endogenous pathways via Notch’s mechanistically direct signaling pathway. We therefore asked if we could satisfy the remaining criteria—programming sensitivity to input forces—by creating novel NRR domains with distinct mechanical properties, which could be used to specify new tension requirements for signaling.

(A) Schematic of tension-mediated activation in Notch or SynNotch receptors. Application of sufficient tensile force via the LBD activates the receptor by displacing three LIN12-Notch repeat (LNR) modules and converting the NRR into a substrate for proteolysis at S2. Cleavage at S2, and concomitantly at S3, liberates the ICD. (LNR modules green; S2 and S3 red) (B) Activating the Notch1 NRR (left) requires tensile force to disrupt the intramolecular interactions that promote an autoinhibited conformation. Engineered sNRR domains (right) include an intramolecularly bound scFv for additional stability. The heterodimerization domain and LNR’s of the NRR are blue and green, respectively, and the scFv added in sNRR is magenta. Molecular interactions between the LNR’s and HD (left) or scFv and NRR (right) are yellow. (PDB 3ETO and 3L95.) (C) Immunostaining of NRR- and sNRR-based receptors for surface-receptor with anti-myc (green) and available NRR with exogenous soluble anti-NRR scFv (magenta) in HeLa cells. Scale bar 25 μm. (D) Schematic of TGT assay used to evaluate molecular tension needed to activate engineered receptors. Fluorescein (FITC) is used as a ligand for SynNotch receptors expressing an anti-FITC scFv LBD. (E) NRR- or sNRR-based receptors with a Gal4-VP64 ICD induce expression of a UAS-controlled H2B-mCherry reporter upon activation. HEK293FT cells transiently expressing these receptors were cultured on TGT-coated surfaces, and reporter activity was monitored by flow cytometry. (F) Tension-induced myogenic differentiation. C3H/10T1/2 fibroblasts express NRR- or sNRR-based SynNotch receptors with a tTA ICD. Upon activation, these receptors drive expression of p65-MyoD, which in turn leads to differentiation. (G) Immunostaining identifies differentiation into skeletal myocytes by multinucleation and positive myosin heavy chain (MHC) expression (green). Scale bar 25 μm.
We began by designing a synthetic NRR domain that required increased tension levels to activate. Single molecule force spectroscopy has shown that proteins can be mechanically stabilized via the binding of other proteins domains (14, 25), and studies of the NRR are consistent with these observations. The binding of antibodies can impart stability upon the NRR, elevating its resistance against chemical- and force-induced unfolding in vitro (20, 26) and limiting the spontaneous activation of mutant receptors in vivo (27, 28). Given these observations, we hypothesized that the NRR’s mechanical stability could be increased by direct fusion with a NRR-binding protein. Specifically, we fused the NRR with a single chain variable fragment (scFv) derived from a NRR-stabilizing antibody (27), such that the scFv could bind intramolecularly to the NRR in its closed state (Fig. 1B). In this way the integrated scFv can impart a stabilizing effect on the NRR, and we anticipated that SynNotch receptors containing these “strengthened NRR” (sNRR) domains would require greater tension to activate.
Engineered mechanoreceptors preserve functional properties of their predecessors
We first sought to verify that sNRR-based receptors preserved the fundamental trafficking and signaling functionalities of their NRR-based predecessors. The receptors tested differed only in the presence of a NRR or sNRR mechanosensing domain, having identical LBD and ICD components. As expected, both NRR- and sNRR-containing SynNotch receptors were processed by the Golgi-localized furin convertase, and the resulting heterodimers were successfully trafficked to the cell surface (Fig. 1C, S1A-B). Labeling of live cells with soluble ligand indicated that the synthetic receptors were constitutively internalized from the cell surface (Fig S1C), similar to the known recycling of natural Notch from the plasma membrane (29). Next, we demonstrated that integration of the anti-NRR scFv blocked sNRR-SynNotch reactivity against soluble anti-NRR, providing evidence that the fused antibody fragment interacts with the NRR epitope in the intended intramolecular manner (Fig. 1C, S1B-C). In addition to synthetic receptors, full-length Notch1 receptors maintained the desired expression properties upon substitution of the NRR for sNRR (Fig. S1D-E).
We used cell-based analyses with a luciferase reporter to confirm that sNRR-SynNotch could be activated by ligand-mediated tension, as delivered by surface-adsorbed ligand. We chose fluorescein dye (FITC) as a ligand to stimulate receptors expressing an anti-FITC LBD, due to the molecule’s well-characterized binding properties, biocompatibility, and versatility. Treatment with either BB-94 or DAPT inhibited signaling, indicating that the activation of sNRR-SynNotch involves proteolysis by a metalloproteinase and ɣ-secretase, respectively (Fig. S2A). Treatment with soluble ligand did not induce luciferase upregulation in NRR- or sNRR-SynNotch expressing cells, consistent with prior reports (24, 30); soluble ligand is unable to offer tensile resistance and instead competitively inhibited activation, further reinforcing the mechanical requirement for receptor activation mechanism (Fig. S2B). Together, these results confirmed that the processing and activation of sNRR-SynNotch proceed in a manner resembling that of both its native and synthetic predecessors.
Engineered receptors require greater tension to activate
Having confirmed the functional similarities between NRR- and sNRR-based SynNotch receptors, we next determined the tensile force required to activate each domain using the tension gauge tether (TGT) assay. In this approach, short double stranded DNA (dsDNA) sequences of known tension tolerance (Ttol) are used to attach ligands (here, FITC) to a culture surface via biotin-streptavidin interaction prior to adding cells (Fig. 1D) (21). These dsDNA tethers (“TGTs”) impose a ceiling on the magnitude of tension that can be applied to individual ligand-bound receptors. If the tension needed to activate sNRR-SynNotch surpasses the Ttol of a given TGT, then the tether will dissociate prior to inducing receptor signaling. If instead the Ttol of a TGT surpasses what is needed to unravel the sNRR, then the tether is expected to endure and stimulate receptor signaling. We used biotin connected to FITC via a non-rupturable linker, rather than via dsDNA, as an upper limit for ligand Ttol, as biotin-streptavidin will tolerate in excess of 100 pN before dissociating (31).
Previous studies have narrowed the activation force of the NRR to 4 – 12 pN (20–23), which is an order of magnitude below the critical force for rupturing antibody-antigen pairs (32). We therefore anticipated that sNRR would exhibit a significantly elevated force-activation threshold. By challenging receptor-expressing reporter cells against TGTs of distinct Ttol values, we defined an activation threshold of approximately > 54 pN for engineered sNRR-SynNotch receptors, well above the < 12 pN threshold known for the Notch1 NRR (Fig. 1E). Other existing NRR-binding antibodies were fused as scFv’s to the NRR to evaluate the generality of our design, and multiple increased receptor tension tolerance (28, 33) (Fig. S3). These data demonstrate that sNRR-SynNotch receptors have increased force requirements for activation since they respond differentially to molecular tensions even while binding the same ligand.
Engineering tension-induced differentiation
Equipped with two mechanically distinct SynNotch receptors, we next aimed to specify how a cell responds to a specific force value. In natural systems, cells enact intricate biological outputs based upon mechanical information in their microenvironment. For example, mechanical forces can guide formation of sarcomeres (34), coordinate wound healing (35), or induce differentiation (7). Using a synthetic biology approach, we generated mouse embryonic fibroblasts that differentiate in response to molecular tension. To do so, we rendered MyoD, a master regulator of myogenic differentiation, SynNotch-dependent. A p65-MyoD fusion under control of the TRE promoter was stably integrated into C3H/10T1/2 fibroblasts (36). SynNotch receptors bearing TetR-VP64 ICD’s were expressed in these cells such that signal transduction would lead to p65-MyoD expression and subsequently to myogenic conversion (Fig. 1F). Using these engineered fibroblasts, we then compared the ability of NRR-SynNotch and sNRR-SynNotch to guide cell fate decisions in response to mechanically distinct TGTs. We observed that cells expressing the receptors differentiated and fused into multinucleated skeletal myocytes only in response to tensions greater than the threshold defined by their respective mechanosensing domains—at least 12 pN for the NRR, and at least 54 pN for sNRR (Fig. 1G, S4).
Systematically generating mechanically distinct receptor isoforms
Further intricacy in cellular mechanosensation can arise from protein isoforms that have distinct mechanical properties despite overlapping biochemical functions, as studied with integrins or talins (37, 38). We anticipated that an expanded collection of sNRR isoforms with diversified force activation thresholds would similarly enable more sophisticated signaling networks. Mutating residues involved in non-covalent interactions at a protein’s “mechano-active” site can lead to distinct mechanical phenotypes (39). We hypothesized that point mutations within the scFv:NRR interface would alter the mechanical stability of sNRR, providing a basis to systematically tune force activation thresholds (Fig. 2A). Using a structure-guided strategy, we generated single- and double-mutants by introducing conservative and non-conservative amino acid substitutions within the scFv:NRR interface (Fig. 2B-C).
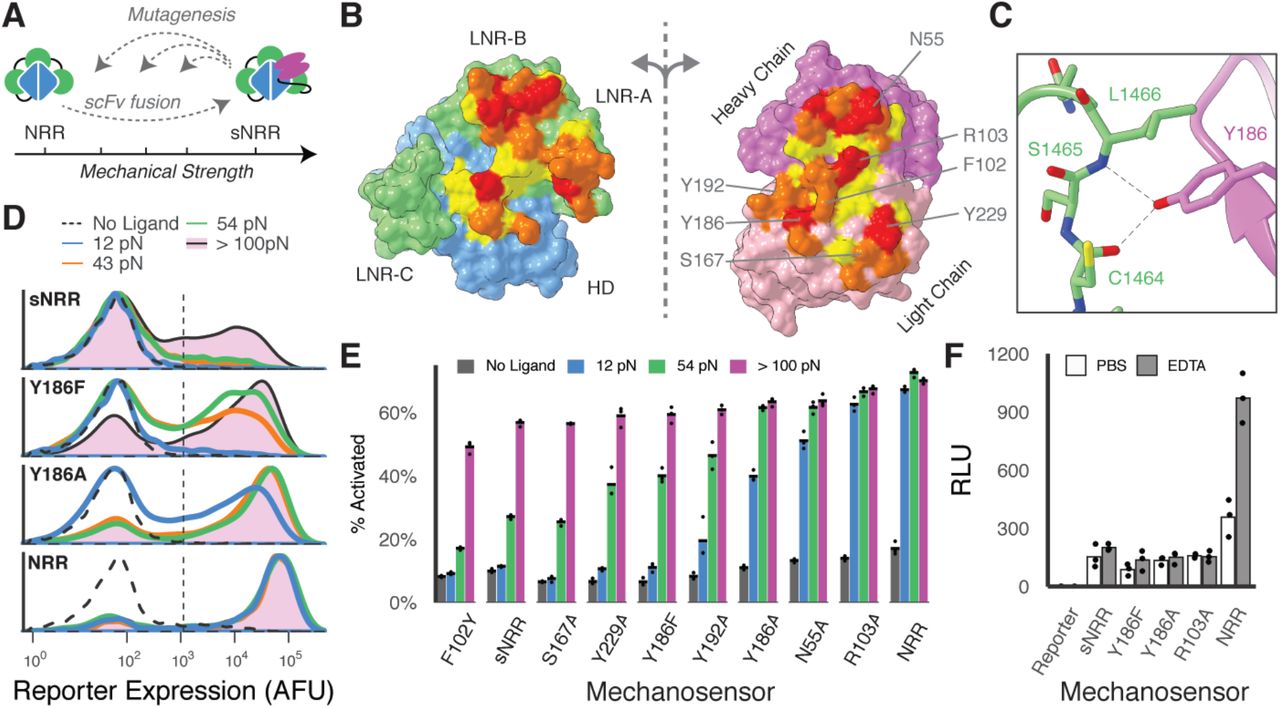
(A) Schematic of strategy to tune mechanical strength. (B) Open-book view of the interface between the Notch1 NRR and the NRR-binding Fab. The HD and LNR’s of the NRR are blue and green, respectively, and the heavy and light chains of the scFv are magenta and pink. Additional residue coloring corresponds to the distance between the two surfaces (< 5.5 A, yellow; 5.5 – 4 A, orange; 4 – 3 A, red) (PDB 3L95). (C) Y186 on the Fab light chain is highlighted as an example mutated residue. (D) Mutating Y186 decreases sNRR mechanical strength. Receptors stably integrated in HEK293FT cells upregulate UAS-driven H2B-mCherry reporter upon activation with TGT’s. (E) Mutating sNRR residues creates receptors with a range of force activation thresholds. Receptors are transiently expressed in HEK293FT cells with stably integrated UAS-H2B-mCherry reporter. Darker shading of bars denotes the extent to which a given TGT activated receptor signaling beyond that of the preceding weaker stimulus. (F) Cells expressing SynNotch receptors with an NRR domain or sNRR domain of various strengths are treated with either 0.5 mM EDTA or PBS for 30 minutes at 37°C, then treatment was quenched with culture media. Reporter expression was evaluated 6 hours later. Activated receptors upregulate expression of a UAS-regulated luciferase.
Consistent with our expectation, introducing mutations resulted in domains of distinct strength, as quantified using a fluorescent reporter gene and the TGT assay (Fig. 2D-E). Mutation generally lead to sNRR isoforms with lower activation thresholds, creating domains with a spectrum of tension tolerances that span a range of biologically relevant forces (40) and populate the activation space between NRR and sNRR. For example, conservatively mutating Tyr186 to Phe decreased the activation threshold below 54 pN, and more severely mutating to Ala furthered this effect. Weakening single mutations were generally additive when combined into doubly mutated domains (Fig. S5A). One mutation (F102Y) may have led to an increase in mechanical stability, though marginal and difficult to resolve using TGT’s (Fig. S5B-C). Of particular use was the Y186F mutation (sNRRY186F), which is weakened such that it activates to a greater extent than the original sNRR on strong TGT’s while still resisting activation by 12 pN TGT’s. Immunostaining confirmed that mutated receptors were trafficked to the cell surface with comparable efficiencies (Fig. S6). We also note that sNRR-based receptors exhibit reduced ligand-independent activation (LIA) in the absence of stimulus, as compared to their NRR-based counterpart (Fig. 2E). Reduced LIA has been targeted as a means to improve SynNotch functionality (41). The difference in LIA for sNRR is increasingly apparent when the ICD is replaced with more potent transcriptional activators VP64-p65 or VP64-p65-rtTA (Fig. S7).
Further analysis of sNRR mechanical strength showed that scFv insertion provides the primary mechanism of stabilizing these engineered domains. In T-cell acute lymphoblastic leukemia (T-ALL), mutations within the NRR can result in destabilized domains that readily undergo LIA (42). However, these same T-ALL mutations, when distal to the scFv binding site, did not affect the mechanical strength of sNRR. Instead, presence of the scFv rescues the hyperactive phenotype, providing evidence that scFv insertion provides stabilization that is a new rate-limiting step in receptor activation (Fig. S8). Only NRR mutations at the scFv binding interface destabilized sNRR-SynNotch receptors, consistent with results from our collection of scFv mutations. NRR mechanical stability depends on the presence of calcium ions, and LIA can be ectopically induced through calcium chelation by EDTA (43). Interestingly, sNRR domains resist activation by EDTA treatment (Fig. 2F). This result holds true for mechanically weakened sNRR variants, further indicating that the scFv fusion provides a dominant autoinhibitory effect. As EDTA is a routine reagent used in cell culture, sNRR domains serendipitously provide a SynNotch variant that can be stably integrated and expanded in cells while resisting inadvertent LIA.
Synthetic mechanogenetic circuits specify how cells interpret mechanical forces
Using our collection of mechanosensitive receptors, we demonstrated further versatility in cellular outputs by designing gene circuits that selectively filter various magnitudes of tension. Synthetic and natural signaling networks can use filtering logic to detect an input, such as small molecule concentration, only if it falls within a desired range. Described in this framework, the mechanogenetic circuits shown thus far generate “high-pass filters,” in which a target gene is expressed only in response to tension beyond a threshold Ttol. Recognizing the engineering utility of “low-pass” and “band-pass” filters that detect low and intermediate forces, respectively, we wished to extend our sNRR-based genetic circuits to achieve these complex outputs.
To build a low-pass filter, we devised a circuit in which a target gene is expressed only if tensions fall below an activation threshold (Fig. S9A). In this approach, activation of a sNRRY186F-SynNotch receptor directs the transcription of a microRNA (miRNA), which in turn inhibits the translation of a constitutively-transcribed fluorescent protein (mCerulean) (Fig. S9B). We observe that this circuit downregulates mCerulean fluorescence in response to increasing molecular tension, as designed. This behavior is reminiscent of natural mechanisms in which mesenchymal stem cells use miRNA to downregulate gene expression in response to stiff ECM (44). In addition to the transcriptional systems demonstrated thus far, the use of miRNA also highlights the ability to use force-activated post-transcriptional control.
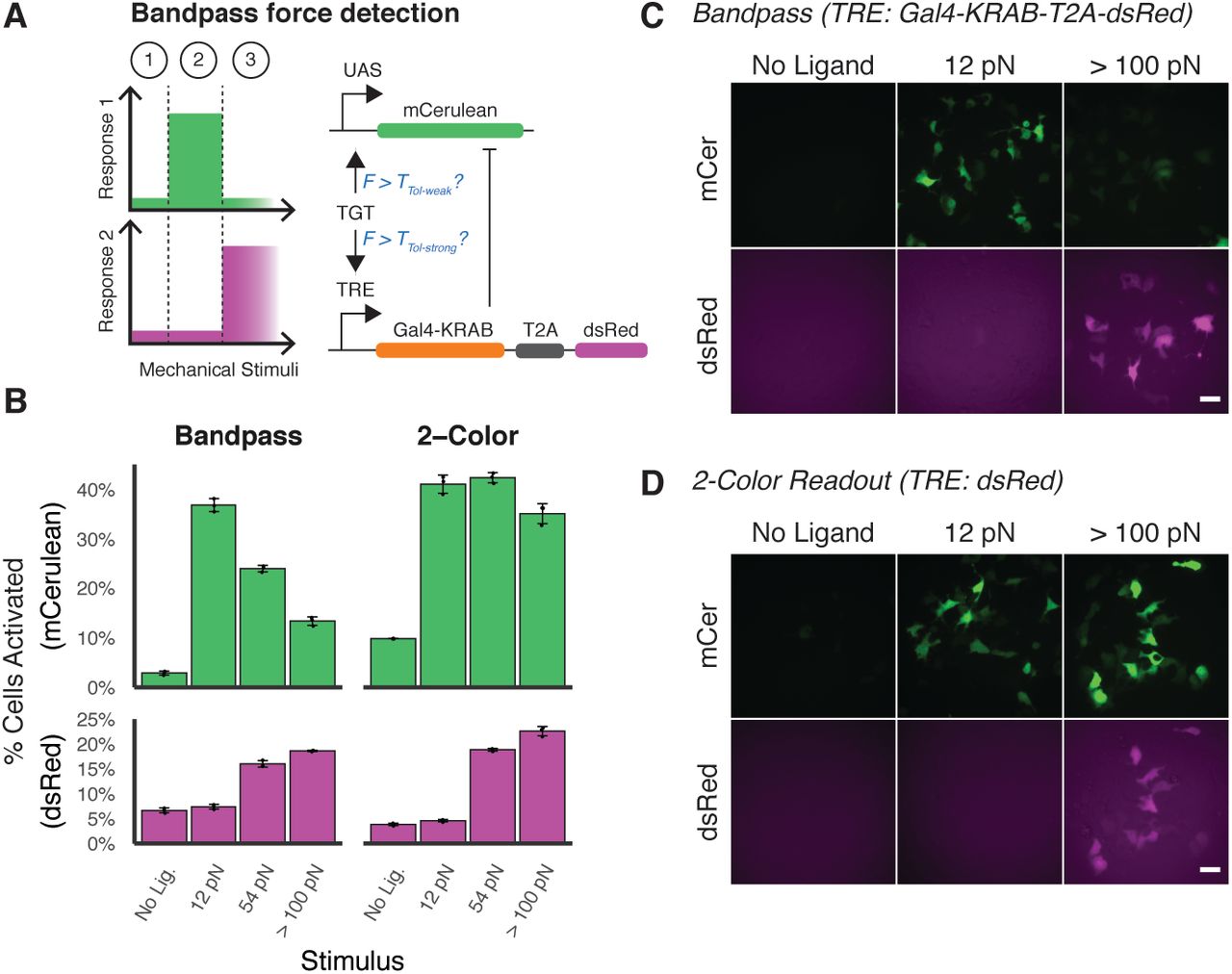
Finally, we engineered a band-pass filter that detects intermediate force magnitudes. A band-pass behavior could more closely mimic that of induced pluripotent stem cells (iPSCs), which pursue distinct cell fates in response to soft, medium, or rigid ECM stiffnesses (7). To achieve this, we combined multiple SynNotch proteins in a single cell to implement an incoherent feedforward loop (IFFL) (Fig. 3A). The IFFL is a signaling motif commonly employed to create a band-pass response with respect to time or concentration (45, 46), and we hypothesized that we could extend the IFFL to similarly create a band-pass behavior with respect to molecular tension. In the activating arm of the loop, an NRR-containing SynNotch possessing a Gal4-VP64 ICD is used to induce mCerulean expression. In the repressing arm of the loop, a sNRRY186F-based receptor containing a tTA ICD is used to drive expression of a Gal4-KRAB fusion protein, which is able to repress expression of mCerulean. Co-transfected cells encoding the IFFL circuit exhibited the expected response profiles, with mCerulean expression occurring only in response to intermediate tension values (Fig. 3B-D). Cells transfected with a control circuit that does not express a Gal4-KRAB inhibitor instead produce a two-color readout of molecular tension.

(A) An IFFL is designed to respond to intermediate forces. A NRR-based SynNotch with Gal4-VP64 ICD drives mCerulean reporter activity upon activation. A sNRRY186F-based SynNotch with tTA ICD drives dsRed, as well as Gal4-KRAB to inhibit mCerulean production. (B) mCerulean (top) and dsRed (bottom) activation of mechanogenetic circuits. “Bandpass” (left) denotes cells expressing the full bandpass circuit in (A). “2-Color” (right) denotes cells expressing this circuit without the Gal4-KRAB inhibitory component. (C-D) Fluorescence images of reporter activity in HEK293FT cells expressing Bandpass or 2-Color readout circuits. Scale bar 25 μm.
In summary, by building upon synthetic Notch proteins, we have designed receptors that are modular in terms of mechanical activation threshold and output transcriptional activity, with minimal crosstalk to other signaling pathways. sNRR and its mutants provide a “kit” of genetically encoded tensiometers that decipher mechanical forces and record them as biochemical responses, providing a quantifiable readout of a cell’s current or past mechanical environment. The generalized nature of this platform enables force-induced control over many processes including genetic recombination and cellular metabolism. This platform also has potential applications in tissue or animal models, where readouts from technologies such as FRET tension sensors may not be viable. Engineered mechanosensitive pathways enable an additional layer of sophistication in synthetic signaling networks, as mechanical forces can propagate information on greater length-scales and faster time-scales than diffusing small molecules. Our work demonstrates the power of structural data in guiding cellular engineering. As more mechanosensitive pathways are elucidated, we anticipate additional proteins will be harnessed to detect the diverse mechanical stimuli that cells experience—tension, compression, shear, and elasticity, both in intra- and extracellular contexts. Continuing to integrate proteins into the mechanogenetic “toolkit” will be critical in empowering us to manipulate mechanobiology in contexts of basic research, targeted therapeutics, and tissue engineering.
Acknowledgments
D.C.S. was supported by an NSF Graduate Research Fellowship. Funding for this work was provided by the NIH (R35 GM128859).





{kind=link}
{kind=link}
{kind=link}